La génétique des populations est une branche de la génétique qui étudie les différences génétiques au sein des populations et entre elles , et fait partie de la biologie évolutive . Les études menées dans cette branche de la biologie examinent des phénomènes tels que l'adaptation , la spéciation et la structure des populations .
La génétique des populations a joué un rôle essentiel dans l' émergence de la synthèse évolutive moderne . Ses principaux fondateurs furent Sewall Wright , J.B.S. Haldane et Ronald Fisher , qui ont également posé les bases de la génétique quantitative, discipline connexe . Traditionnellement très mathématique, la génétique des populations moderne englobe des travaux théoriques, de laboratoire et de terrain. Les modèles de génétique des populations sont utilisés à la fois pour l'inférence statistique à partir de données de séquences d'ADN et pour la validation ou l'invalidation de concepts.
Ce qui distingue la génétique des populations des approches plus récentes et phénotypiques de modélisation de l'évolution, telles que la théorie des jeux évolutionnaires et la dynamique adaptative , c'est l'importance qu'elle accorde à des phénomènes génétiques comme la dominance , l'épistasie , le degré de rupture du déséquilibre de liaison par la recombinaison génétique , ainsi que les phénomènes aléatoires de mutation et de dérive génétique . C'est ce qui la rend pertinente pour la comparaison avec les données de la génomique des populations .
Histoire
La génétique des populations est née de la volonté de concilier l'hérédité mendélienne et les modèles biostatistiques . La sélection naturelle n'entraîne l'évolution que s'il existe une variation génétique suffisante au sein d'une population. Avant la découverte de la génétique mendélienne , l'une des hypothèses courantes était celle de l'hérédité par mélange . Cependant, avec ce modèle, la variance génétique serait rapidement perdue, rendant l'évolution par sélection naturelle ou sexuelle peu plausible. Le principe de Hardy-Weinberg explique comment la variation est maintenue dans une population à hérédité mendélienne. Selon ce principe, les fréquences alléliques (variations d'un gène) restent constantes en l'absence de sélection, de mutation, de migration et de dérive génétique.


L'étape suivante, cruciale, fut le travail du biologiste et statisticien britannique Ronald Fisher . Dans une série d'articles, débutant en 1918 et culminant avec son ouvrage de 1930, *The Genetical Theory of Natural Selection* , Fisher démontra que la variation continue mesurée par les biométriciens pouvait être produite par l'action combinée de nombreux gènes discrets, et que la sélection naturelle pouvait modifier les fréquences alléliques au sein d'une population, engendrant ainsi l'évolution. Dans une série d'articles commençant en 1924, un autre généticien britannique, J.B.S. Haldane , élabora les modèles mathématiques de la variation de fréquence allélique au niveau d'un locus génique unique dans un large éventail de conditions. Haldane appliqua également l'analyse statistique à des exemples concrets de sélection naturelle, tels que l'évolution de la phalène du bouleau et le mélanisme industriel , et montra que les coefficients de sélection pouvaient être supérieurs à ceux supposés par Fisher, conduisant à une évolution adaptative plus rapide, notamment comme stratégie de camouflage face à une pollution accrue.
Le biologiste américain Sewall Wright , spécialiste des expériences de reproduction animale , s'est intéressé aux interactions génétiques et aux effets de la consanguinité sur de petites populations relativement isolées présentant une dérive génétique. En 1932, il a introduit le concept de paysage adaptatif et a soutenu que la dérive génétique et la consanguinité pouvaient éloigner une petite sous-population isolée d'un pic adaptatif, permettant ainsi à la sélection naturelle de la diriger vers d'autres pics adaptatifs.
Les travaux de Fisher, Haldane et Wright ont fondé la génétique des populations. Cette discipline a intégré la sélection naturelle à la génétique mendélienne, constituant ainsi une première étape cruciale vers l'élaboration d'une théorie unifiée de l'évolution. John Maynard Smith fut l'élève de Haldane, tandis que W.D. Hamilton fut influencé par les écrits de Fisher. L'Américain George R. Price collabora avec Hamilton et Maynard Smith. L'Américain Richard Lewontin et le Japonais Motoo Kimura furent influencés par Wright et Haldane.
Synthèse moderne
Les mathématiques de la génétique des populations ont été initialement développées comme point de départ de la synthèse moderne . Des auteurs tels que Beatty ont affirmé que la génétique des populations constitue le cœur de cette synthèse. Durant les premières décennies du XXe siècle, la plupart des naturalistes de terrain continuaient de croire que le lamarckisme et l'orthogenèse offraient la meilleure explication de la complexité qu'ils observaient dans le monde vivant . Lors de la synthèse moderne, ces idées ont été écartées et seules les causes évolutives pouvant être exprimées dans le cadre mathématique de la génétique des populations ont été conservées . Un consensus a été atteint quant aux facteurs évolutifs susceptibles d'influencer l'évolution, mais pas quant à l'importance relative de ces différents facteurs
Theodosius Dobzhansky , chercheur postdoctoral au sein du laboratoire de T.H. Morgan , fut influencé par les travaux sur la diversité génétique de généticiens russes tels que Sergueï Tchetverikov . Avec son ouvrage de 1937, *Génétique et origine des espèces* , il contribua à combler le fossé entre les fondements de la microévolution développés par les généticiens des populations et les modèles de macroévolution observés par les biologistes de terrain. Dobzhansky examina la diversité génétique des populations sauvages et démontra que, contrairement aux hypothèses des généticiens des populations, ces populations présentaient une grande diversité génétique, avec des différences marquées entre les sous-populations. L'ouvrage vulgarisa également les travaux très mathématiques des généticiens des populations. Grâce à Dobzhansky, la génétique des populations fut influencée par un nombre bien plus important de biologistes que ceux qui avaient accès aux ouvrages originaux, très techniques.
En Grande-Bretagne, E.B. Ford , pionnier de la génétique écologique a continué, tout au long des années 1930 et 1940, à démontrer empiriquement le pouvoir de la sélection naturelle exercée par des facteurs écologiques, notamment la capacité à maintenir la diversité génétique grâce à des polymorphismes génétiques tels que les groupes sanguins humains . Les travaux de Ford, menés en collaboration avec Fisher, ont contribué à un changement d'orientation lors de la synthèse moderne, qui a mis en avant la sélection naturelle comme force dominante.
Théorie neutre et dynamique de fixation de l'origine
La conception synthétique moderne originelle de la génétique des populations postule que les mutations constituent une matière première abondante et se concentre uniquement sur la variation de fréquence des allèles au sein des populations . Les principaux processus influençant les fréquences alléliques sont la sélection naturelle , la dérive génétique , le flux génique et les mutations récurrentes . Fisher et Wright ont présenté des désaccords fondamentaux quant aux rôles respectifs de la sélection et de la dérive . La disponibilité de données moléculaires sur toutes les différences génétiques a conduit à la théorie neutre de l'évolution moléculaire . Selon cette conception, de nombreuses mutations sont délétères et ne sont donc jamais observées, tandis que la plupart des autres sont neutres, c'est-à-dire qu'elles ne sont pas soumises à la sélection. Le destin de chaque mutation neutre étant laissé au hasard (dérive génétique), la direction du changement évolutif est déterminée par les mutations qui surviennent et ne peut donc être expliquée par les seuls modèles de variation de fréquence des allèles (existants)
La vision de l'origine-fixation de la génétique des populations généralise cette approche au-delà des mutations strictement neutres et considère le taux auquel un changement particulier se produit comme le produit du taux de mutation et de la probabilité de fixation .
Quatre processus
Sélection
La sélection naturelle , qui inclut la sélection sexuelle , correspond au fait que certains caractères augmentent les chances de survie et de reproduction d'un organisme . La génétique des populations décrit la sélection naturelle en définissant la valeur sélective comme une propension ou une probabilité de survie et de reproduction dans un environnement donné. Cette valeur sélective est généralement exprimée par le symbole w = 1 - s, où s est le coefficient de sélection . La sélection naturelle agissant sur les phénotypes , les modèles de génétique des populations supposent des relations relativement simples pour prédire le phénotype, et donc la valeur sélective, à partir de l' allèle présent à un ou quelques loci. Ainsi, la sélection naturelle transforme les différences de valeur sélective entre individus présentant des phénotypes différents en modifications de la fréquence allélique au sein d'une population, au fil des générations.
Avant l'avènement de la génétique des populations, de nombreux biologistes doutaient que de faibles différences de valeur sélective suffisent à influencer significativement l'évolution. Les généticiens des populations ont abordé cette question en partie en comparant la sélection à la dérive génétique . La sélection peut compenser la dérive génétique lorsque s est supérieur à 1 divisé par la taille effective de la population . Lorsque ce critère est rempli, la probabilité qu'un nouveau mutant avantageux se fixe est approximativement égale à 2s . Le temps nécessaire à la fixation d'un tel allèle est approximativement de 10⁻⁵ .
Dominance
La dominance signifie que l’effet phénotypique et/ou adaptatif d’un allèle à un locus dépend de l’allèle présent dans la seconde copie de ce locus. Considérons trois génotypes à un locus, avec les valeurs adaptatives suivantes
| Génotype: | A 1 A 1 | A 1 A 2 | A 2 A 2 |
| Forme physique relative : | 1 | 1 heure | 1-s |
s est le coefficient de sélection et h le coefficient de dominance. La valeur de h fournit les informations suivantes :
| h=0 | A1 dominant , A2 récessif |
| h=1 | A2 dominant , A1 récessif |
| 0<h<1 | dominance incomplète |
| h<0 | dominance excessive |
| h>1 | Sous-dominance |
Épistase

L'épistasie signifie que l'effet phénotypique et/ou adaptatif d'un allèle à un locus donné dépend des allèles présents à d'autres locus. La sélection n'agit pas sur un seul locus, mais sur un phénotype résultant du développement d'un génotype complet. Cependant, de nombreux modèles de génétique des populations d'espèces sexuées sont des modèles « à un seul locus », où l'adaptation d'un individu est calculée comme le produit des contributions de chacun de ses locus, ce qui revient à supposer l'absence d'épistasie.
En réalité, la relation entre génotype et valeur sélective est plus complexe. La génétique des populations doit soit modéliser cette complexité en détail, soit la saisir par une règle moyenne plus simple. Empiriquement, les mutations bénéfiques tendent à avoir un impact moindre sur la valeur sélective lorsqu'elles sont introduites dans un contexte génétique déjà fortement sélectif : c'est ce qu'on appelle l'épistasie à rendements décroissants . Lorsque les mutations délétères ont également un effet moindre sur la valeur sélective dans des contextes génétiques fortement sélectifs, on parle d'« épistasie synergique ». Cependant, l'effet des mutations délétères tend en moyenne à être très proche d'un effet multiplicatif, voire peut présenter le schéma inverse, appelé « épistasie antagoniste »
L’épistasie synergique est centrale dans certaines théories de l’élimination de la charge mutationnelle et dans l’ évolution de la reproduction sexuée .
Mutation

Le processus de mutation génétique se déroule chez un individu et entraîne des modifications héréditaires du matériel génétique. Ce processus est souvent caractérisé par la description des états initial et final, ou du type de modification survenue au niveau de l'ADN (par exemple, une mutation T→C, une délétion d'un nucléotide), des gènes ou des protéines (par exemple, une mutation nulle, une mutation de perte de fonction), ou à un niveau phénotypique supérieur (par exemple, la mutation responsable des yeux rouges). Les mutations mononucléotidiques sont fréquemment les plus courantes, mais de nombreux autres types de mutations sont possibles et surviennent à des taux très variables, pouvant présenter des asymétries ou des biais systématiques ( biais mutationnels ).
Les mutations peuvent impliquer la duplication de larges portions d'ADN , généralement par recombinaison génétique . Ceci entraîne une variation du nombre de copies au sein d'une population. Les duplications constituent une source majeure de matériel génétique pour l'évolution de nouveaux gènes . D'autres types de mutations créent parfois de nouveaux gènes à partir d'ADN non codant
Dans la distribution des effets sur la valeur sélective (DFE) des nouvelles mutations, seule une minorité d'entre elles sont bénéfiques. Les mutations ayant des effets importants sont généralement délétères. Des études menées sur la drosophile (Drosophila melanogaster) suggèrent que si une mutation modifie une protéine produite par un gène, cela sera probablement nocif, environ 70 % de ces mutations ayant des effets néfastes, les autres étant soit neutres, soit faiblement bénéfiques.
Ce processus biologique de mutation est représenté dans les modèles de génétique des populations de deux manières : soit comme une pression déterministe de mutations récurrentes sur les fréquences alléliques, soit comme une source de variation. Dans la théorie déterministe, l’évolution débute avec un ensemble prédéterminé d’allèles et se poursuit par des variations continues de leurs fréquences, comme si la population était infinie. L’apparition de mutations chez les individus est représentée par une « force » ou une « pression » de mutation à l’échelle de la population, c’est-à-dire la force d’innombrables mutations d’intensité u appliquée aux variations de fréquence f(A1) à f(A2). Par exemple, dans le modèle classique d’équilibre mutation-sélection , la pression de mutation augmente la fréquence d’un allèle, tandis que la sélection contre ses effets délétères la diminue, jusqu’à atteindre un équilibre donné (dans le cas le plus simple) par f = u/s.
Ce concept de pression mutationnelle est surtout utile pour examiner les implications des mutations délétères, telles que la charge mutationnelle et ses conséquences sur l'évolution du taux de mutation. La transformation des populations par la pression mutationnelle est improbable. Haldane a avancé qu'elle nécessiterait des taux de mutation élevés sans opposition de la sélection, et Kimura a conclu, de manière encore plus pessimiste, que même cela était improbable, car le processus serait trop long (voir l'évolution par pression mutationnelle ).
Cependant, l'évolution par pression mutationnelle est possible dans certaines circonstances et est depuis longtemps considérée comme une cause possible de la perte de caractères non utilisés . Par exemple, les pigments deviennent inutiles chez les animaux vivant dans l'obscurité des grottes et tendent à disparaître . Un exemple expérimental concerne la perte de sporulation dans des populations expérimentales de *B. subtilis* . La sporulation est un caractère complexe codé par de nombreux loci, de sorte que le taux de mutation associé à sa perte a été estimé à une valeur exceptionnellement élevée . Dans ce cas, la perte de sporulation peut survenir par mutation récurrente, sans nécessiter de sélection pour la perte de cette capacité. En l'absence de sélection pour la perte de fonction, la vitesse d'évolution de cette perte dépend davantage du taux de mutation que de la taille effective de la population , ce qui indique qu'elle est davantage déterminée par la mutation que par la dérive génétique.
Le rôle de la mutation comme source de nouveauté diffère des modèles classiques de pression mutationnelle. Lorsque les modèles de génétique des populations intègrent un processus d'introduction ou d'apparition de mutations dépendant du taux, c'est-à-dire un processus introduisant de nouveaux allèles, y compris neutres et bénéfiques, les propriétés de la mutation peuvent avoir un impact plus direct sur le rythme et la direction de l'évolution, même à faible taux de mutation. Autrement dit, le spectre des mutations peut devenir crucial, notamment les biais mutationnels (différences prévisibles dans les taux d'occurrence des différents types de mutations), car un biais dans l'introduction de la variation peut influencer le cours de l'évolution.
La mutation joue un rôle clé dans d'autres théories classiques et récentes, notamment le cliquet de Muller , la subfonctionnalisation , le concept de catastrophe d'erreur d'Eigen et l'hypothèse du risque mutationnel de Lynch .
Dérive génétique
La dérive génétique est une modification des fréquences alléliques due à un échantillonnage aléatoire . Autrement dit, les allèles présents chez la descendance constituent un échantillon aléatoire de ceux présents chez les parents . La dérive génétique peut entraîner la disparition complète de variants génétiques et, par conséquent, réduire la variabilité génétique. Contrairement à la sélection naturelle, qui rend les variants génétiques plus ou moins fréquents en fonction de leur succès reproductif les modifications dues à la dérive génétique ne sont pas induites par des pressions environnementales ou adaptatives et ont autant de chances de rendre un allèle plus fréquent que moins fréquent.
L'effet de la dérive génétique est plus important pour les allèles présents en peu de copies que pour ceux présents en grand nombre. La génétique des populations, et notamment la dérive génétique, est décrite soit par des processus de ramification , soit par une équation de diffusion décrivant les variations de fréquence allélique . Ces approches sont généralement appliquées aux modèles de Wright-Fisher et de Moran . En supposant que la dérive génétique soit la seule force évolutive agissant sur un allèle, après t générations dans de nombreuses populations répliquées, avec des fréquences alléliques initiales de p et q, la variance de la fréquence allélique entre ces populations est :
Ronald Fisher considérait que la dérive génétique ne jouait qu'un rôle mineur dans l'évolution, et cette opinion est restée dominante pendant plusieurs décennies. Aucune perspective de génétique des populations n'a jamais accordé à la dérive génétique un rôle central à elle seule, mais certaines ont mis en évidence son importance en combinaison avec une autre force non sélective. La théorie de l'équilibre fluctuant de Sewall Wright affirmait que la combinaison de la structure des populations et de la dérive génétique était importante. La théorie neutre de l'évolution moléculaire de Motoo Kimura soutient que la plupart des différences génétiques au sein des populations et entre elles sont dues à la combinaison de mutations neutres et de dérive génétique.
Le rôle de la dérive génétique par échantillonnage dans l'évolution a été critiqué par John H. Gillespie et Will Provine [ soutiennent que la sélection sur des sites liés constitue une force stochastique plus importante, accomplissant le travail traditionnellement attribué à la dérive génétique par échantillonnage. Les propriétés mathématiques de la dérive génétique diffèrent de celles de la dérive génétique . La direction de la variation aléatoire de la fréquence allélique est autocorrélée d'une génération à l'autre
flux génétique
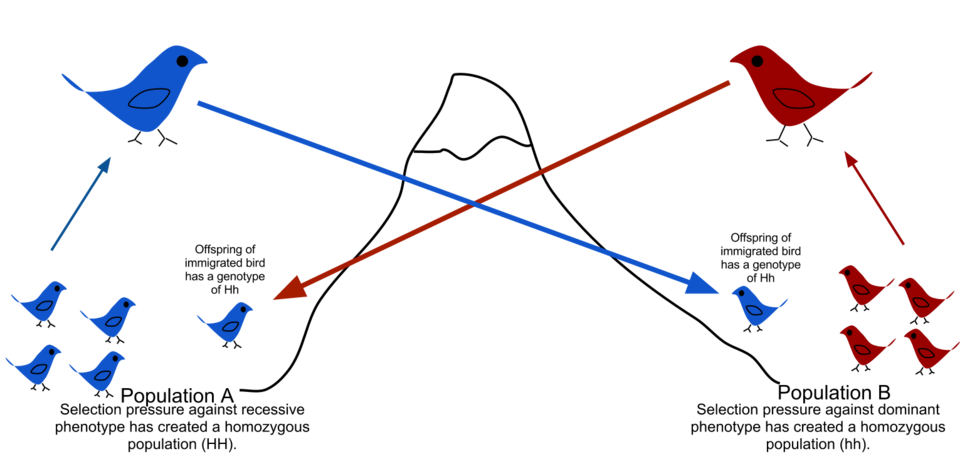
En raison des barrières physiques à la migration, de la faible propension des individus à se déplacer ou à se disperser ( vagilité ) et de leur tendance à rester ou à retourner à leur lieu de naissance ( philopatrie ), les populations naturelles se reproduisent rarement entre elles, contrairement à ce que pourraient supposer certains modèles aléatoires théoriques ( panmixie ) . Il existe généralement une aire de répartition géographique au sein de laquelle les individus sont plus étroitement apparentés que ceux sélectionnés aléatoirement dans la population générale. Ce phénomène est décrit comme le degré de structuration génétique d'une population

La structuration génétique peut être causée par des migrations dues à des changements climatiques historiques , à l'expansion de l'aire de répartition des espèces ou à la disponibilité actuelle d' habitats . Le flux génétique est entravé par les chaînes de montagnes, les océans et les déserts, voire par des structures artificielles telles que la Grande Muraille de Chine , qui a freiné la circulation des gènes végétaux.
Le flux génique est l'échange de gènes entre populations ou espèces, entraînant une rupture de leur structure. Au sein d'une même espèce, le flux génique peut se manifester par la migration puis la reproduction d'organismes, ou encore par l'échange de pollen . Le transfert de gènes entre espèces inclut la formation d' organismes hybrides et le transfert horizontal de gènes . Les modèles de génétique des populations permettent d'identifier les populations présentant un isolement génétique significatif et de reconstituer leur histoire.
L’isolement d’une population entraîne une dépression de consanguinité . Les migrations au sein d’une population peuvent introduire de nouvelles variantes génétiques , contribuant potentiellement à un sauvetage évolutif . Si une proportion significative d’individus ou de gamètes migrent, cela peut également modifier les fréquences alléliques, par exemple en engendrant une charge migratoire .
En présence de flux génétique, d'autres barrières à l'hybridation entre deux populations divergentes d'une espèce exogame sont nécessaires pour que ces populations deviennent de nouvelles espèces .
transfert horizontal de gènes
Le transfert horizontal de gènes est le transfert de matériel génétique d'un organisme à un autre qui n'est pas sa descendance ; ce phénomène est très fréquent chez les procaryotes . En médecine, il contribue à la propagation de la résistance aux antibiotiques , car lorsqu'une bactérie acquiert des gènes de résistance, elle peut rapidement les transférer à d'autres espèces . Des transferts horizontaux de gènes de bactéries vers des eucaryotes, tels que la levure Saccharomyces cerevisiae et le bruche du haricot azuki Callosobruchus chinensis, ont également pu se produire . Les rotifères bdelloïdes eucaryotes , qui semblent avoir reçu divers gènes de bactéries, de champignons et de plantes, constituent un exemple de transferts à plus grande échelle . Les virus peuvent également transporter de l'ADN entre organismes, permettant ainsi le transfert de gènes même entre différents domaines biologiques . Un transfert de gènes à grande échelle s'est également produit entre les ancêtres des cellules eucaryotes et des procaryotes, lors de l'acquisition des chloroplastes et des mitochondries .
Lien
Si tous les gènes sont en équilibre de liaison , l'effet d'un allèle à un locus peut être moyenné sur l'ensemble du pool génique aux autres loci. En réalité, un allèle est fréquemment en déséquilibre de liaison avec des gènes situés à d'autres loci, notamment avec des gènes voisins sur le même chromosome. La recombinaison rompt ce déséquilibre de liaison trop lentement pour éviter l'effet de « trajet génétique indirect », où un allèle à un locus atteint une fréquence élevée parce qu'il est lié à un allèle soumis à la sélection à un locus voisin. La liaison ralentit également le taux d'adaptation, même dans les populations sexuées. L'effet du déséquilibre de liaison sur le ralentissement du taux d'évolution adaptative résulte d'une combinaison de l' effet Hill-Robertson (retards dans le rapprochement des mutations bénéfiques) et de la sélection de fond (retards dans la séparation des mutations bénéfiques des mutations délétères transmises par un « trajet génétique indirect »).
La liaison génétique pose problème aux modèles de génétique des populations qui traitent un locus génique à la fois. Elle peut toutefois être exploitée comme méthode pour détecter l'action de la sélection naturelle par le biais de balayages sélectifs .
Dans le cas extrême d'une population asexuée , la liaison génétique est complète et les équations de génétique des populations peuvent être dérivées et résolues en termes d' onde progressive des fréquences génotypiques le long d'un paysage adaptatif simple . La plupart des microbes , comme les bactéries , sont asexués. La génétique des populations de leur adaptation présente deux régimes contrastés. Lorsque le produit du taux de mutation bénéfique et de la taille de la population est faible, les populations asexuées suivent un « régime de succession » de dynamique d'origine-fixation, le taux d'adaptation étant fortement dépendant de ce produit. Lorsque ce produit est beaucoup plus élevé, les populations asexuées suivent un régime de « mutations concurrentes », le taux d'adaptation étant moins dépendant du produit et caractérisé par l'interférence clonale et l'apparition d'une nouvelle mutation bénéfique avant que la précédente ne soit fixée .
Applications
Explication des niveaux de variation génétique
La théorie neutre prédit que le niveau de diversité nucléotidique au sein d'une population est proportionnel au produit de la taille de la population et du taux de mutation neutre. Le fait que les niveaux de diversité génétique varient beaucoup moins que la taille des populations est connu sous le nom de « paradoxe de la variation » . Si une forte diversité génétique constituait l'un des arguments initiaux en faveur de la théorie neutre, le paradoxe de la variation est devenu l'un des arguments les plus convaincants contre cette théorie.
Il est clair que les niveaux de diversité génétique varient considérablement au sein d'une espèce en fonction du taux de recombinaison local, en raison à la fois de l'effet de sélection indirecte et de la sélection de fond . La plupart des solutions actuelles au paradoxe de la variation invoquent un certain niveau de sélection sur des sites liés. Par exemple, une analyse suggère que les populations plus importantes subissent davantage de balayages sélectifs, ce qui élimine une plus grande diversité génétique neutre. Une corrélation négative entre le taux de mutation et la taille de la population peut également contribuer à ce phénomène.
L’histoire de vie influence davantage la diversité génétique que l’histoire des populations, par exemple les espèces à stratégie r présentent une plus grande diversité génétique.
Détection de la sélection
Les modèles de génétique des populations permettent de déterminer quels gènes sont soumis à la sélection. Une approche courante consiste à rechercher des régions de fort déséquilibre de liaison et de faible variance génétique le long du chromosome, afin de détecter des balayages sélectifs récents .
Une autre approche courante est le test de McDonald-Kreitman , qui compare la variation au sein d'une espèce ( polymorphisme ) à la divergence entre espèces (substitutions) sur deux types de sites ; l'un d'eux étant considéré comme neutre. Généralement, les sites synonymes sont considérés comme neutres. Les gènes soumis à une sélection positive présentent un excès de sites divergents par rapport aux sites polymorphes. Ce test permet également d'obtenir une estimation à l'échelle du génome de la proportion de substitutions fixées par la sélection positive, α. Selon la théorie neutre de l'évolution moléculaire , cette proportion devrait être proche de zéro. Des valeurs élevées ont donc été interprétées comme une réfutation à l'échelle du génome de la théorie neutre.
Inférence démographique
Le test le plus simple pour détecter une structure de population chez une espèce diploïde à reproduction sexuée consiste à vérifier si les fréquences génotypiques suivent les proportions de Hardy-Weinberg en fonction des fréquences alléliques. Par exemple, dans le cas le plus simple d'un seul locus avec deux allèles , notés A et a , de fréquences respectives p et q , l'appariement aléatoire prédit freq( AA ) = p /2 pour les homozygotes AA , freq( aa ) = q /2 pour les homozygotes aa et freq( Aa ) = 2pq pour les hétérozygotes . En l'absence de structure de population, les proportions de Hardy-Weinberg sont atteintes en une à deux générations d'appariement aléatoire. Plus généralement, on observe un excès d'homozygotes, signe de structure de population. L'importance de cet excès peut être quantifiée par le coefficient de consanguinité, F.
Les individus peuvent être regroupés en K sous-populations. Le degré de structuration de la population peut ensuite être calculé à l'aide de F<sub> ST</sub> , qui mesure la proportion de variance génétique expliquée par la structure de la population. La structure génétique de la population peut alors être mise en relation avec la structure géographique, et un mélange génétique peut être détecté.
La théorie de la coalescence relie la diversité génétique d'un échantillon à l'histoire démographique de la population dont il est issu. Elle suppose généralement la neutralité , et par conséquent, les séquences provenant de régions évoluant de manière plus neutre des génomes sont sélectionnées pour de telles analyses. Elle permet d'inférer les relations entre espèces ( phylogénétique ), ainsi que la structure des populations, leur histoire démographique (par exemple, les goulots d'étranglement démographiques , la croissance démographique ), la dispersion biologique , la dynamique source-puits et l'introgression au sein d'une espèce.
Une autre approche de l’inférence démographique repose sur le spectre de fréquence des allèles .
Évolution des systèmes génétiques
En supposant l'existence de loci contrôlant le système génétique lui-même, des modèles de génétique des populations sont élaborés pour décrire l' évolution de la dominance et d'autres formes de robustesse , l' évolution de la reproduction sexuée et des taux de recombinaison, l'évolution des taux de mutation , l'évolution des mécanismes de régulation évolutive , l'évolution des caractères de signalisation coûteux , l' évolution du vieillissement et l'évolution de la coopération . Par exemple, la plupart des mutations étant délétères, le taux de mutation optimal pour une espèce peut résulter d'un compromis entre les dommages causés par un taux élevé de mutations délétères et les coûts métaboliques liés au maintien de systèmes visant à réduire ce taux, tels que les enzymes de réparation de l'ADN.
Un aspect important de ces modèles est que la sélection n'est suffisamment forte pour éliminer les mutations délétères et, par conséquent, contrer le biais mutationnel vers la dégradation que si le coefficient de sélection s est supérieur à l'inverse de la taille effective de la population . Ce phénomène est connu sous le nom de barrière de dérive génétique et est lié à la théorie quasi neutre de l'évolution moléculaire . La théorie de la barrière de dérive génétique prédit que les espèces à grande taille de population effective posséderont des systèmes génétiques très rationalisés et efficaces, tandis que celles à petite taille de population auront des génomes volumineux et complexes contenant par exemple des introns et des éléments transposables . Cependant, de manière quelque peu paradoxale, les espèces à grande taille de population pourraient être si tolérantes aux conséquences de certains types d'erreurs qu'elles développent des taux d'erreur plus élevés, par exemple en transcription et en traduction , que les petites populations