En génétique des populations , le flux génique (également appelé migration ou flux allélique ) désigne le transfert de matériel génétique d'une population à une autre. Si le taux de flux génique est suffisamment élevé, deux populations présenteront des fréquences alléliques équivalentes et pourront donc être considérées comme une seule population effective. Il a été démontré qu'un seul migrant par génération suffit à empêcher la divergence des populations due à la dérive génétique . Les populations peuvent diverger sous l'effet de la sélection , même lors d'échanges d'allèles, si la pression de sélection est suffisamment forte Le flux génique est un mécanisme important de transfert de diversité génétique entre les populations. Les migrants modifient la distribution de la diversité génétique au sein des populations en modifiant les fréquences alléliques (la proportion d'individus porteurs d'une variante particulière d'un gène). Des taux élevés de flux génique peuvent réduire la différenciation génétique entre deux groupes, augmentant ainsi leur homogénéité . On a longtemps pensé que le flux génique limitait la spéciation et prévenait l'expansion des aires de répartition en combinant les pools génétiques des groupes, empêchant ainsi le développement de différences de variation génétique qui auraient conduit à la différenciation et à l'adaptation . Dans certains cas, la dispersion entraînant un flux de gènes peut également entraîner l'ajout de nouvelles variantes génétiques sous sélection positive au pool génétique d'une espèce ou d'une population (introgression adaptative. )
Plusieurs facteurs influencent le flux génétique entre différentes populations. Ce flux est généralement plus faible chez les espèces peu mobiles ou à faible dispersion, vivant dans des habitats fragmentés, lorsque les distances entre les populations sont importantes et lorsque les populations sont de petite taille. La mobilité joue un rôle crucial dans le taux de dispersion, les individus très mobiles ayant généralement de meilleures perspectives de déplacement. Bien que les animaux soient considérés comme plus mobiles que les plantes, le pollen et les graines peuvent être transportés sur de grandes distances par les animaux, l'eau ou le vent. Lorsque le flux génétique est entravé, la consanguinité peut augmenter , comme l'indique le coefficient de consanguinité (F) au sein d'une population. Par exemple, de nombreuses populations insulaires présentent de faibles taux de flux génétique en raison de leur isolement géographique et de leur petite taille. Le wallaby des rochers à pieds noirs compte plusieurs populations consanguines vivant sur différentes îles au large des côtes australiennes. L'isolement de cette population est tel que l'absence de flux génétique a entraîné des taux élevés de consanguinité.
taille effective de la population (σ ) et du taux de migration net par génération (m). En utilisant l'approximation basée sur le modèle insulaire, l'effet de la migration peut être calculé pour une population en termes de degré de différenciation génétique (ΔG ). Cette formule tient compte de la proportion de la variation totale des marqueurs moléculaires entre les populations, moyennée sur les loci . Lorsqu'il y a un migrant par génération, ΔG est égal à 0,2. Cependant, lorsqu'il y a moins d'un migrant par génération (absence de migration), le coefficient de consanguinité augmente rapidement, entraînant une fixation et une divergence complète (β = 1). La valeur la plus courante est inférieure à 0,25, ce qui indique une certaine migration. Les mesures de la structure des populations varient de 0 à 1. Lorsque des flux de gènes se produisent par migration, les effets néfastes de la consanguinité peuvent être atténués.Obstacles au flux génétique
Spéciation allopatrique
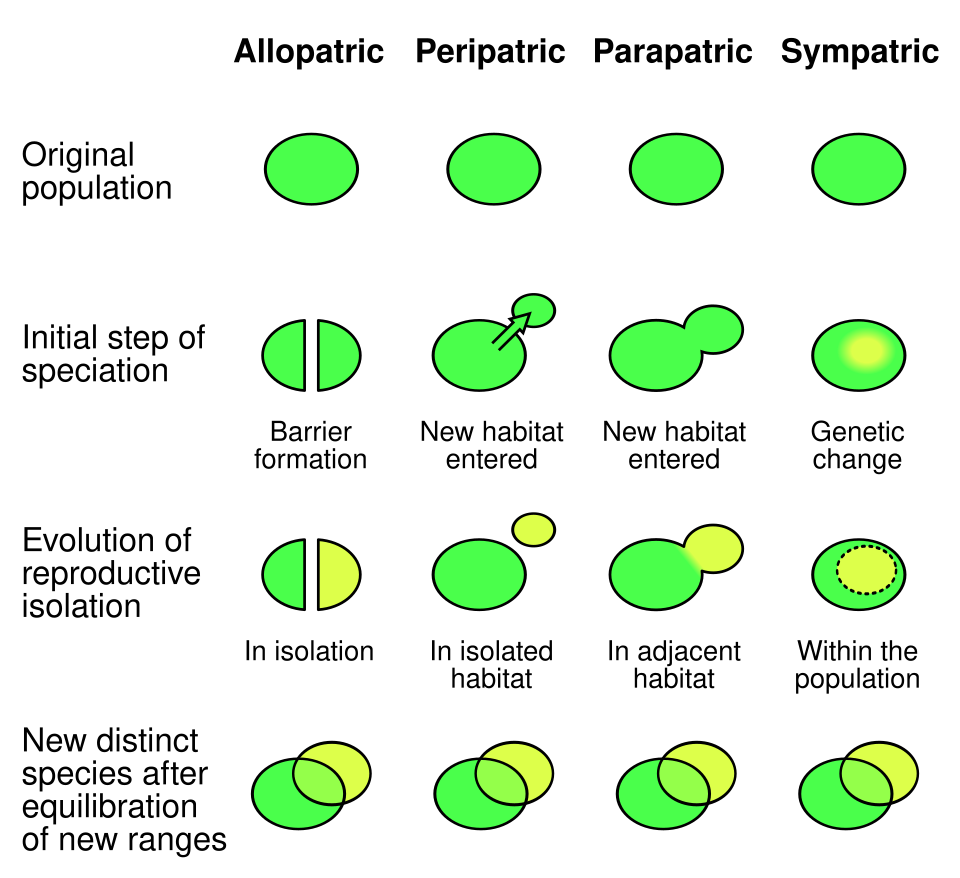
Lorsque les échanges génétiques sont bloqués par des barrières physiques, il en résulte une spéciation allopatrique , c'est-à-dire un isolement géographique qui empêche les populations d'une même espèce d'échanger du matériel génétique. Ces barrières sont généralement naturelles, comme les chaînes de montagnes infranchissables, les océans ou les vastes déserts. Cependant, elles peuvent parfois être artificielles, à l'instar de la Grande Muraille de Chine , qui a entravé les échanges génétiques de populations végétales indigènes. L'une de ces plantes indigènes, l'orme de Chine (Ulmus pumila) , présente une prévalence de différenciation génétique plus faible que les plantes Vitex negundo, Ziziphus jujuba, Heteropappus hispidus et Prunus armeniaca , dont l'habitat se situe de l'autre côté de la Grande Muraille . Ceci s'explique par le fait que l'orme de Chine se propage principalement par pollinisation anémophile , tandis que les autres plantes sont pollinisées par les insectes. Il a été démontré que des échantillons de la même espèce qui poussent de part et d’autre ont développé des différences génétiques, car il y a peu ou pas de flux de gènes pour assurer la recombinaison des pools génétiques.pollution génétique , accouplement assortatif et exogamie . Chez l'humain, la différenciation génétique peut également résulter de l'endogamie , due aux différences de caste, d'appartenance ethnique, de coutumes et de religion.
Flux de gènes assisté par l'homme
Sauvetage génétique
Le flux génique peut également être utilisé pour aider les espèces menacées d'extinction. Lorsqu'une espèce vit en petites populations, le risque de consanguinité est accru et la vulnérabilité à la perte de diversité due à la dérive génétique est plus grande. Ces populations peuvent grandement bénéficier de l'introduction d'individus non apparentés , capables d'accroître la diversité , de réduire la consanguinité et, potentiellement, d'augmenter la taille de la population . Ceci a été démontré en laboratoire avec deux souches de drosophile (Drosophila melanogaster ) présentant un goulot d'étranglement génétique : des croisements entre les deux populations ont inversé les effets de la consanguinité et ont permis d'améliorer les chances de survie non seulement sur une génération, mais sur deux
flux de gènes assisté
À l'instar du sauvetage génétique, le flux génique assisté est une stratégie de gestion permettant de conserver les espèces menacées en simulant les processus naturels de flux génique. En déplaçant des individus ou des gamètes entre populations au sein de l'aire de répartition actuelle d'une espèce, le flux génique assisté contribue à atténuer les effets du changement climatique en facilitant l'adaptation aux nouvelles conditions climatiques locales. Il permet d'introduire des génotypes préadaptés aux conditions climatiques souhaitées ou d'accroître la fréquence des génotypes déjà adaptés, améliorant ainsi la valeur sélective globale des populations confrontées à un changement climatique rapide. Bien que cette stratégie puisse être utilisée pour sauver des espèces subissant un goulot d'étranglement démographique, ses applications concernent toute espèce dont l'habitat sera affecté par des changements environnementaux importants.
pollution génétique
La facilitation urbaine peut se manifester de diverses manières, mais la plupart des mécanismes impliquent la mise en contact, directe ou indirecte, d'espèces auparavant séparées. L'urbanisation, en modifiant un habitat, entraîne sa fragmentation, mais peut aussi potentiellement perturber les barrières et créer un corridor reliant deux espèces autrefois isolées. L'efficacité de ce processus dépend des capacités de dispersion et d'adaptation de chaque espèce à différents environnements, ainsi que de sa capacité à utiliser les infrastructures anthropiques pour se déplacer. Le changement climatique d'origine humaine est un autre mécanisme par lequel les animaux vivant dans les régions méridionales peuvent être contraints de migrer vers le nord, vers des températures plus fraîches, où ils pourraient entrer en contact avec d'autres populations absentes de leur aire de répartition. Plus directement, l'introduction d'espèces non indigènes dans de nouveaux environnements par l'homme peut conduire à l' hybridation d'espèces similaires.
Ce modèle de facilitation urbaine a été testé sur un parasite nuisible pour la santé humaine, la veuve noire de l'Ouest ( Latrodectus hesperus ). Une étude de Miles et al. a recueilli des données de variation des polymorphismes nucléotidiques simples (SNP) à l'échelle du génome chez des populations d'araignées urbaines et rurales. Les résultats ont mis en évidence un flux génique accru chez les veuves noires de l'Ouest urbaines par rapport aux populations rurales. De plus, le génome de ces araignées était plus similaire entre les populations rurales qu'entre les populations urbaines, suggérant une diversité accrue, et donc une meilleure adaptation, chez les populations urbaines de la veuve noire de l'Ouest. Phénotypiquement, les araignées urbaines sont plus grandes, plus foncées et plus agressives, ce qui pourrait expliquer leur meilleure survie en milieu urbain. Ces résultats appuient l'hypothèse de la facilitation urbaine, car ces araignées sont effectivement capables de se propager et de se diversifier plus rapidement en milieu urbain qu'en milieu rural. Cependant, cela illustre également comment la facilitation urbaine, malgré l'augmentation du flux génique, n'est pas nécessairement bénéfique pour l'environnement, car le venin de la veuve noire de l'Ouest est très toxique et représente donc un risque pour la santé humaine.
Un autre exemple de facilitation urbaine est celui des migrations du lynx roux ( Lynx rufus ) dans le nord des États-Unis et le sud du Canada. Une étude menée par Marrote et al. a séquencé quatorze loci microsatellites différents chez des lynx roux de la région des Grands Lacs et a révélé que la longitude influençait l'interaction entre les modifications anthropiques du paysage et le flux génétique au sein de la population. Si la hausse des températures mondiales pousse les populations de lynx roux vers le nord, l'intensification des activités humaines favorise également leur migration vers cette région. Cette intensification entraîne une augmentation du réseau routier et de la circulation, mais aussi une augmentation de l'entretien des routes, du déneigement et du compactage de la neige, créant ainsi involontairement un passage pour les lynx roux. L'influence anthropique sur les voies de migration du lynx roux illustre la facilitation urbaine par l'ouverture d'un corridor de flux génétique. Cependant, dans la partie sud de l'aire de répartition du lynx roux, l'augmentation du réseau routier et de la circulation est corrélée à une diminution du couvert forestier, ce qui entrave le flux génétique au sein de la population. Paradoxalement, le déplacement des lynx roux vers le nord est causé par le réchauffement climatique d'origine humaine, mais est également facilité par l'augmentation de l'activité anthropique dans les zones septentrionales qui rendent ces habitats plus adaptés aux lynx roux.
Les conséquences de la facilitation urbaine varient d'une espèce à l'autre. La facilitation urbaine peut avoir des effets positifs lorsque l'augmentation du flux génétique permet une meilleure adaptation et introduit des allèles bénéfiques, ce qui, idéalement, accroîtrait la biodiversité. Ceci a des implications pour la conservation : par exemple, la facilitation urbaine profite à une espèce de tarentule menacée et pourrait contribuer à augmenter la taille de sa population. Des effets négatifs se produisent lorsque l'augmentation du flux génétique est inadaptée et entraîne la perte d'allèles bénéfiques. Dans le pire des cas, cela conduirait à une extinction génomique par le biais d'un essaim hybride . Il est également important de noter que, dans le contexte de la santé globale des écosystèmes et de la biodiversité, la facilitation urbaine n'est pas nécessairement bénéfique et s'applique généralement aux ravageurs adaptés aux milieux urbains. On peut citer comme exemples la veuve noire de l'Ouest mentionnée précédemment, ainsi que le crapaud buffle , qui a pu utiliser les routes pour se déplacer et proliférer en Australie.
flux génétique entre espèces
transfert horizontal de gènes
Les virus peuvent transférer des gènes entre espèces. Les bactéries peuvent incorporer des gènes provenant de bactéries mortes, échanger des gènes avec des bactéries vivantes et échanger des plasmides au-delà des frontières d'espèces. « Les comparaisons de séquences suggèrent un transfert horizontal récent de nombreux gènes entre diverses espèces , y compris au-delà des frontières des « domaines » phylogénétiques . Ainsi, la détermination de l'histoire phylogénétique d'une espèce ne peut être réalisée de manière concluante en déterminant les arbres évolutifs de gènes isolés. »
Le biologiste Gogarten suggère que « la métaphore originale de l’arbre ne correspond plus aux données issues des recherches récentes sur le génome ». Les biologistes devraient plutôt utiliser la métaphore de la mosaïque pour décrire les différentes histoires combinées dans les génomes individuels et celle d’un réseau entrelacé pour visualiser la richesse des échanges et les effets coopératifs du transfert horizontal de gènes.
« L’utilisation de gènes uniques comme marqueurs phylogénétiques rend difficile la reconstitution de la phylogénie des organismes en présence de transferts horizontaux de gènes (THG). La combinaison du modèle simple de coalescence de la cladogenèse avec de rares événements de THG suggère l’absence d’un ancêtre commun unique ayant contenu tous les gènes ancestraux partagés par les trois domaines du vivant . Chaque molécule contemporaine possède sa propre histoire et remonte à un ancêtre moléculaire individuel . Cependant, ces ancêtres moléculaires étaient probablement présents chez différents organismes à différentes époques. »
Hybridation
Dans certains cas, lorsqu'une espèce possède une espèce sœur et que la reproduction est possible grâce à la levée d'anciennes barrières ou à l'introduction d'espèces par l'homme, les espèces peuvent s'hybrider et échanger des gènes et des caractères associés. Cet échange n'est pas toujours évident : les hybrides peuvent parfois être phénotypiquement identiques à l'espèce d'origine , mais l'analyse de l'ADNmt révèle une hybridation. L'hybridation différentielle se produit également car certains caractères et l'ADN s'échangent plus facilement que d'autres, sous l'effet de la pression de sélection ou de son absence, ce qui facilite les échanges. Lorsque l'espèce introduite commence à remplacer l'espèce indigène, cette dernière est menacée et la biodiversité diminue, faisant de ce phénomène un exemple négatif plutôt qu'un exemple positif de flux génique augmentant la diversité génétique. L'introgression est le remplacement des allèles d'une espèce par ceux de l'espèce envahissante. Les hybrides sont parfois moins « aptes » que leur génération parentale, et constituent par conséquent un problème génétique étroitement surveillé, car l’objectif ultime de la génétique de la conservation est de maintenir l’intégrité génétique d’une espèce et de préserver la biodiversité.
Exemples

Bien que les flux génétiques puissent considérablement améliorer la valeur sélective d'une population, ils peuvent aussi avoir des conséquences négatives selon la population et son environnement. Les effets des flux génétiques dépendent du contexte.
- Population fragmentée : les paysages fragmentés comme celui des îles Galápagos constituent un lieu idéal pour la radiation adaptative, résultant de la diversité géographique. Les pinsons de Darwin ont probablement subi une spéciation allopatrique, en partie due à cette diversité géographique, mais cela n’explique pas la présence de tant d’espèces différentes de pinsons sur une même île. Ce phénomène est dû à la radiation adaptative, c’est-à-dire l’évolution de caractères variés en réponse à la compétition pour les ressources. Le flux génétique s’oriente vers les ressources abondantes à un moment donné.
- Population insulaire : L’ iguane marin est une espèce endémique des îles Galápagos, mais il descend d’un ancêtre continental de l’iguane terrestre. En raison de l’isolement géographique, les échanges génétiques entre les deux espèces ont été limités et les environnements différents ont contraint l’iguane marin à évoluer pour s’adapter au milieu insulaire. Par exemple, c’est le seul iguane à avoir développé la capacité de nager.
- Populations humaines : En Europe, Homo sapiens s'est croisé avec les Néandertaliens, entraînant un flux génétique entre ces populations. Ce flux génétique a conduit à la présence d'allèles néandertaliens dans la population européenne moderne. Deux théories expliquent l' évolution humaine à travers le monde. La première, dite multirégionale, considère la variation humaine moderne comme le produit de la radiation d' Homo erectus hors d'Afrique, suivie d'une différenciation locale menant à l'établissement des populations régionales telles que nous les connaissons aujourd'hui. Le flux génétique joue un rôle important dans le maintien d'un certain degré de similarité et la prévention de la spéciation. À l'inverse, la théorie de l'origine unique postule l'existence d'une population ancestrale commune d' Homo sapiens, originaire d'Afrique, qui présentait déjà les caractéristiques anatomiques actuelles. Cette théorie minimise l'importance de l'évolution parallèle nécessaire.
- Papillons : Des comparaisons entre les populations sympatriques et allopatriques d’ Heliconius melpomene , H. cydno et H. timareta ont révélé une tendance à l’échelle du génome d’une variation partagée accrue en sympatrie, indiquant un flux de gènes interspécifique omniprésent.
- Flux génétiques induits par l'homme : La gestion génétique en captivité des espèces menacées est le seul moyen par lequel l'homme tente d'induire des flux génétiques ex situ. Le panda géant en est un exemple : il fait partie d'un programme international de reproduction où du matériel génétique est partagé entre les organisations zoologiques afin d'accroître la diversité génétique au sein des petites populations. Face à un faible succès reproductif, l'insémination artificielle avec du sperme frais ou congelé-décongelé a été mise au point, ce qui a permis d'augmenter le taux de survie des petits. Une étude de 2014 a estimé que les centres de reproduction présentaient des niveaux élevés de diversité génétique et de faibles niveaux de consanguinité.
- Plantes : Deux populations de mimules ont été trouvées utilisant des pollinisateurs différents (abeilles et colibris) qui ont limité le flux génétique, entraînant un isolement génétique et produisant finalement deux espèces différentes, Mimulus lewisii et Mimulus cardinalis .
- Cerfs sika : Les cerfs sika ont été introduits en Europe occidentale et se reproduisent facilement avec les cerfs élaphes indigènes. Cette translocation a entraîné une introgression génétique, et il n’existe plus de cerfs élaphes « purs » dans la région ; tous peuvent être classés comme hybrides.
- Colin de Virginie : Des colins de Virginie ont été introduits du sud des États-Unis vers l’Ontario afin d’accroître leur population et le gibier destiné à la chasse. Les hybrides issus de cette translocation étaient moins robustes que la population indigène et n’étaient pas adaptés aux hivers rigoureux du Nord.