L'importance commerciale principale des mousses réside dans leur rôle de constituant majeur de la tourbe (principalement le genre Sphagnum ), bien qu'elles soient également utilisées à des fins décoratives, notamment dans les jardins et chez les fleuristes . Traditionnellement, les mousses étaient employées comme isolant et pour leur capacité à absorber jusqu'à 20 fois leur poids en liquide. Espèces clés de voûte , les mousses contribuent à la restauration des habitats et au reboisement . Elles absorbent 6,4 milliards de tonnes de carbone par an et offrent divers autres avantages environnementaux, tels que la lutte contre les agents pathogènes dangereux pour l'homme, la prévention de l'érosion des sols , l'amélioration du microclimat [ la filtration de la pollution atmosphérique .
Du point de vue botanique, les mousses sont des plantes non vasculaires appartenant à l'embranchement des Bryophytes. Ce sont généralement de petites plantes herbacées (non ligneuses) (quelques centimètres de hauteur) qui absorbent l'eau et les nutriments principalement par leurs feuilles et utilisent le dioxyde de carbone et la lumière du soleil pour produire leur propre nourriture par photosynthèse . À l'exception du groupe ancien des Takakiopsida , aucune mousse connue ne forme de mycorhizes , mais les champignons bryphilitiques sont largement répandus chez les mousses et autres bryophytes, où ils vivent en tant que saprophytes, parasites, pathogènes et mutualistes, certains étant des endophytes . Les mousses se distinguent des plantes vasculaires par l'absence de xylème ( trachéides ou vaisseaux) transportant l'eau . Comme chez les hépatiques et les anthocérotes , la génération gamétophytique haploïde est la phase dominante de leur cycle de vie . Cela contraste avec le modèle observé chez les plantes vasculaires ( plantes à graines et ptéridophytes ), où la génération sporophyte diploïde est dominante. Les mousses se reproduisent par spores , et non par graines , et n'ont pas de fleurs.

Les gamétophytes des mousses possèdent des tiges simples ou ramifiées, dressées (acrocarpe) ou prostrées (pleurocarpe). Les classes ayant divergé précocement, Takakiopsida, Sphagnopsida, Andreaeopsida et Andreaeobryopsida, sont soit dépourvues de stomates , soit présentent des pseudo-stomates ne formant pas de pores. Dans les autres classes, les stomates ont disparu plus de 60 fois. Leurs feuilles sont simples, généralement constituées d'une seule couche de cellules sans espaces intercellulaires, et présentent souvent une nervure médiane plus épaisse. Cette nervure peut se prolonger au-delà du bord de l'apex, formant une excroissance. L'extrémité du limbe peut se prolonger en un épiderme composé de cellules incolores, qui apparaissent blanches sur le fond vert foncé de la feuille. Le bord de la feuille peut être lisse ou dentelé. Il peut exister un type cellulaire distinct définissant le bord de la feuille, différant par sa forme et/ou sa couleur des autres cellules foliaires.
Les mousses possèdent des rhizoïdes filiformes qui les ancrent à leur substrat, comparables à des poils absorbants plutôt qu'aux structures racinaires plus importantes des spermatophytes . On sait que les mousses absorbent l'eau par leurs rhizoïdes, et certaines espèces peuvent également absorber des nutriments de cette manière. Elles se distinguent des hépatiques ( Marchantiophyta ou Hepaticae) par leurs rhizoïdes multicellulaires. Les capsules contenant les spores, ou sporanges , des mousses sont portées isolément sur de longues tiges non ramifiées, ce qui les distingue des polysporangiophytes , qui comprennent toutes les plantes vasculaires. Les sporophytes producteurs de spores (c'est-à-dire la génération diploïde multicellulaire) sont généralement capables de photosynthèse, mais ont une durée de vie courte et dépendent du gamétophyte pour leur approvisionnement en eau et la plupart, voire la totalité, de leurs nutriments. De plus, chez la majorité des mousses, la capsule sporifère se développe et arrive à maturité après l'allongement de son pédoncule, tandis que chez les hépatiques, la capsule se développe et arrive à maturité avant l'allongement de son pédoncule. D'autres différences ne sont pas universelles pour toutes les mousses et toutes les hépatiques, mais la présence d'une tige bien différenciée portant des feuilles simples, non vasculaires et non disposées sur trois rangs, indique clairement qu'il s'agit d'une mousse.Les plantes vasculaires possèdent deux jeux de chromosomes dans leurs cellules végétatives et sont dites diploïdes ; chaque chromosome a un homologue contenant la même information génétique, ou une information similaire. En revanche, les mousses et autres bryophytes ne possèdent qu’un seul jeu de chromosomes et sont donc haploïdes (chaque chromosome existe en un seul exemplaire dans la cellule). Il existe une phase, au cours du cycle de vie des mousses, où elles possèdent un double jeu de chromosomes appariés, mais cela se produit uniquement pendant le stade sporophyte .
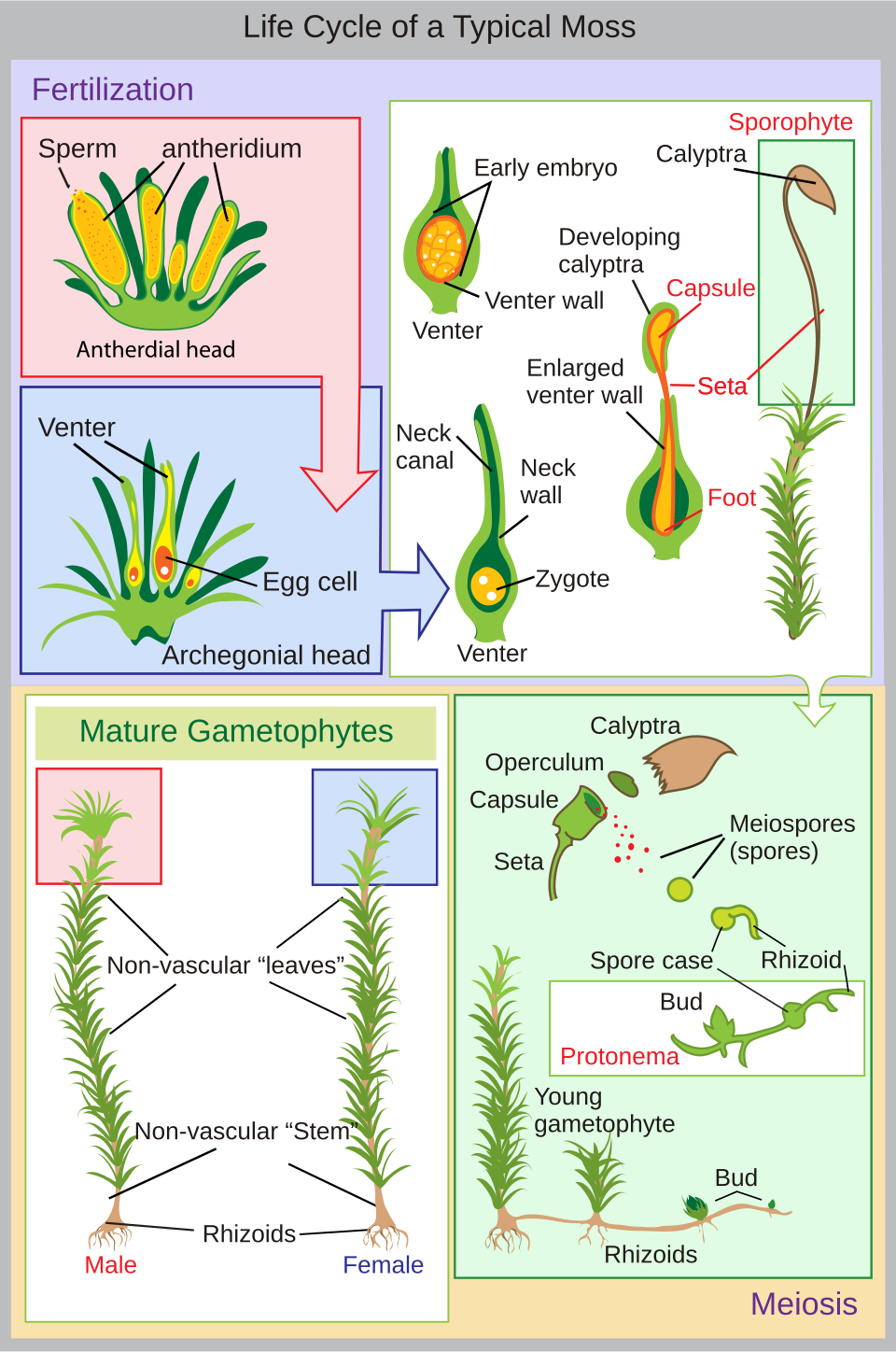
Le cycle de vie des mousses débute par une spore haploïde qui germe pour former un protonéma ( pluriel : protonémata), soit un amas de filaments, soit un thalloïde (plat et en forme de thalle). Les protonémata de mousse, lorsqu'ils sont regroupés, ressemblent généralement à un fin feutre vert et peuvent se développer sur un sol humide, l'écorce d'arbre, les rochers, le béton ou presque toute autre surface relativement stable. Il s'agit d'une étape transitoire dans la vie d'une mousse, mais du protonéma se développe le gamétophore (« porteur de gamètes ») qui se différencie ensuite en tiges et en feuilles. Un seul amas de protonémata peut développer plusieurs pousses gamétophoriques, formant ainsi une touffe de mousse.
À l'extrémité des tiges ou des ramifications du gamétophore se développent les organes reproducteurs des mousses. Les organes femelles, appelés archégones ( singulier : archégone ), sont protégés par un ensemble de feuilles modifiées formant le périchétum (pluriel : périchétes). Les archégones sont de petits amas de cellules en forme de flacon, présentant un col ouvert (ventre) par lequel nagent les spermatozoïdes mâles. Les organes mâles, appelés anthéridies ( singulier : anthéridie ), sont entourés de feuilles modifiées appelées périgonium ( pluriel : périgonies). Chez certaines mousses, les feuilles environnantes forment une cupule, permettant ainsi aux spermatozoïdes qu'elle contient d'être projetés vers les tiges voisines par les gouttelettes d'eau.
La croissance de l'apex du gamétophore est perturbée par la chitine fongique . Galotto et al. (2020) ont utilisé l'expression de leurs gènes . Ils ont conclu que cette réponse de défense était probablement conservée depuis l' ancêtre commun le plus récent des bryophytes et des trachéophytes . Orr et al. (2020) ont découvert que les microtubules des cellules apicales en croissance étaient structurellement similaires à l'actine F et remplissaient une fonction similaire.
Les mousses peuvent être dioïques (à comparer avec les plantes à graines) ou monoïques (à comparer avec les plantes à graines ). Chez les mousses dioïques, les organes reproducteurs mâles et femelles sont portés par des gamétophytes différents. Chez les mousses monoïques (ou autoïques), les deux types d'organes sont portés par le même gamétophyte. En présence d'eau, les spermatozoïdes contenus dans les anthéridies nagent jusqu'aux archégones et la fécondation a lieu, donnant naissance à un sporophyte diploïde. Les spermatozoïdes des mousses sont biflagellés, c'est-à-dire qu'ils possèdent deux flagelles qui facilitent leur propulsion. Puisque les spermatozoïdes doivent nager jusqu'à l'archégone, la fécondation est impossible sans eau. Certaines espèces (par exemple *Mnium hornum* ou plusieurs espèces de *Polytrichum *) conservent leurs anthéridies dans des « coupelles », des structures en forme de bol situées à l'extrémité des tiges, qui propulsent les spermatozoïdes sur plusieurs décimètres lorsque des gouttelettes d'eau les atteignent, augmentant ainsi la distance de fécondation.
Après la fécondation, le sporophyte immature émerge de la face ventrale de l'archégone. Sa maturation prend plusieurs mois . Le sporophyte est composé d'un long pédoncule, appelé soie, et d'une capsule coiffée d'un opercule . La capsule et l'opercule sont eux-mêmes enveloppés par une calyptre haploïde, vestige de la face ventrale de l'archégone. La calyptre se détache généralement à maturité de la capsule. À l'intérieur de celle-ci, les cellules sporifères subissent la méiose pour former des spores haploïdes, permettant ainsi un nouveau cycle. L'ouverture de la capsule est généralement munie d'un péristome, une structure dentelée qui peut être absente chez certaines mousses.genre Sphagnum, les spores sont projetées à environ accélération gravitationnelle terrestre g .

Il a été récemment découvert que les microarthropodes, tels que les collemboles et les acariens , peuvent influencer la fécondation des mousses et que ce processus est modulé par les composés odorants émis par ces dernières. Les mousses de feu mâles et femelles , par exemple, émettent des composés organiques volatils différents et complexes . Les plantes femelles émettent davantage de composés que les plantes mâles. Il a été observé que les collemboles privilégient les plantes femelles, et une étude a montré qu'ils favorisent la fécondation des mousses, suggérant une relation olfactive analogue à la relation plante-pollinisateur observée chez de nombreuses plantes à graines . L'espèce de mousse puante Splachnum sphaericum optimise la pollinisation par les insectes en attirant les mouches vers ses sporanges grâce à une forte odeur de charogne, et en fournissant un signal visuel puissant sous la forme de collerettes rouges renflées sous chaque capsule de spores. Les mouches attirées par la mousse transportent ses spores jusqu'aux excréments frais d'herbivores, qui constituent l'habitat privilégié des espèces de ce genre.
Chez de nombreuses mousses, comme Ulota phyllantha , des structures végétatives vertes appelées propagules se forment sur les feuilles ou les branches. Ces propagules peuvent se détacher et donner naissance à de nouvelles plantes sans fécondation. Il s'agit d'un mode de reproduction asexuée , et ces unités génétiquement identiques peuvent conduire à la formation de populations clonales .
mâles nains
Les mâles nains des mousses (également appelés spores mâles dispersées par le vent qui se déposent et germent sur la tige femelle, où leur croissance est limitée à quelques millimètres. Chez certaines espèces, le nanisme est génétiquement déterminé : toutes les spores mâles deviennent naines . Le plus souvent, il est déterminé par l’environnement : les spores mâles qui se déposent sur une plante femelle deviennent naines, tandis que celles qui se déposent ailleurs se développent en mâles de grande taille, comparables à ceux des femelles . Dans ce dernier cas, les mâles nains transplantés des femelles vers un autre substrat se développent en tiges de grande taille, ce qui suggère que les femelles émettent une substance inhibant la croissance des mâles en germination et accélérant peut-être aussi leur maturation sexuelle . La nature de cette substance est inconnue, mais l’ auxine, une phytohormone, pourrait être impliquée
Le fait que les mâles se développent en tant que individus nains sur les femelles devrait accroître l' efficacité de la fécondation en minimisant la distance entre les organes reproducteurs mâles et femelles. En conséquence, il a été observé que la fréquence de fécondation est positivement corrélée à la présence de mâles nains chez plusieurs espèces lignées non apparentées et pourraient être plus fréquents qu'on ne le pensait auparavant. Par exemple, on estime qu'entre un quart et la moitié de tous les pleurocarpacées dioïques possèdent des mâles nains.
réparation de l'ADN
La mousse *Physcomitrium patens* est utilisée comme organisme modèle pour étudier la réparation des lésions de l'ADN chez les plantes, et plus particulièrement le mécanisme de réparation appelé recombinaison homologue . Si la plante est incapable de réparer les lésions de l'ADN, comme les cassures double brin , dans ses cellules somatiques , ces dernières peuvent perdre leurs fonctions normales ou mourir. Si cela se produit lors de la méiose (étape de la reproduction sexuée), elles peuvent devenir stériles. Le séquençage du génome de *P. patens* a permis d'identifier plusieurs gènes impliqués dans la réparation de l'ADN . Des mutants de *P. patens* présentant des anomalies dans des étapes clés de la recombinaison homologue ont été utilisés pour élucider le fonctionnement de ce mécanisme de réparation chez les plantes. Par exemple, une étude menée sur des mutants de *P. patens* déficients en *Rp RAD51*, un gène codant pour une protéine essentielle à la réaction de réparation par recombinaison, a montré que la recombinaison homologue est indispensable à la réparation des cassures double brin de l'ADN chez cette plante. De même, des études sur des mutants déficients en Ppmre11 ou Pprad50 (qui codent des protéines clés du complexe MRN , le principal capteur des cassures double brin de l'ADN) ont montré que ces gènes sont nécessaires à la réparation des dommages à l'ADN ainsi qu'à la croissance et au développement normaux.
Classification
Depuis 2018, les mousses sont regroupées avec les hépatiques et les anthocérotes dans l'embranchement des Bryophytes ( ou Bryophyta sensu lato ). Cet embranchement comprend trois groupes, parfois considérés comme des embranchements distincts : les Bryophytes (mousses), les Marchantiophytes (hépatiques) et les Anthocérotes (anthocérotes). Il a été proposé de reclasser ces derniers dans les classes des Bryopsides, des Marchantiopsides et des Anthocérotes, respectivement. Les mousses et les hépatiques sont désormais considérées comme appartenant à un clade appelé Setaphyta .
Les mousses (Bryophyta sensu stricto) sont divisées en huit classes :
division Bryophyta
| |
| Bryophytes | |
| Néomusci | |
| Cénomusci | |
| Altamusci | |















