{{The Mollusca|volume=11}} "},"image":{"wt":"Cephalopoda diversity.jpg"},"image_upright":{"wt":"1.2"},"image_caption":{"wt":"Extant and extinct cephalopods; clockwise from top-l...
Worldlex WikiContenu en francaisLecture gratuite
classe mollusques Cephalopoda grec seiche ou tête proéminente un ensemble de bras ou de tentacules ( musculaires à encre », en référence à leur capacité à projeter de l'encre . L'étude des céphalopodes est une branche de la malacologie appelée téuthologie .
Il existe plus de 800 espèces actuelles de céphalopodes , bien que de nouvelles espèces continuent d'être décrites. On estime à 11 000 le nombre de taxons éteints décrits, mais la nature molle des céphalopodes fait qu'ils ne se fossilisent pas facilement
On trouve des céphalopodes dans tous les océans de la Terre. Aucun ne tolère l'eau douce , mais le calmar court, Lolliguncula brevis , présent dans la baie de Chesapeake , constitue une exception partielle notable puisqu'il tolère l'eau saumâtre . On pense que les céphalopodes sont incapables de vivre en eau douce en raison de multiples contraintes biochimiques et, au cours de leurs plus de 400 millions d'années d'existence, ils ne se sont jamais aventurés dans des habitats entièrement d'eau douce.
Les céphalopodes occupent la majeure partie des profondeurs océaniques, des plaines abyssales à la surface, et ont également été trouvés dans la zone hadale . Leur diversité est la plus grande près de l'équateur (environ 40 espèces capturées dans des filets à 11°N lors d'une étude de diversité) et diminue vers les pôles (environ 5 espèces capturées à 60°N).
Biologie
Système nerveux et comportement
À gauche : Une pieuvre ouvrant un récipient à bouchon à vis. À droite : Un calmar bobtail hawaïen ( Euprymna scolopes ) s'enfouissant dans le sable, ne laissant apparaître que ses yeux.
Les céphalopodes sont généralement considérés comme les invertébrés les plus intelligents et possèdent des sens très développés ainsi qu'un cerveau volumineux (plus gros que celui des gastéropodes ) . Leur système nerveux est le plus complexe des invertébrés et leur rapport masse cérébrale/masse corporelle se situe entre celui des vertébrés endothermes et ectothermes . On a observé que des céphalopodes en distance au sol du laboratoire, entrer dans un autre aquarium pour se nourrir de crabes en captivité, puis retourner dans le leur
Le cerveau est protégé par un crâne cartilagineux . Les fibres nerveuses géantes du manteau des céphalopodes sont utilisées depuis de nombreuses années comme matériel expérimental en neurophysiologie ; leur grand diamètre (dû à l’absence de myélinisation ) les rend relativement faciles à étudier par rapport à d’autres animaux.
De nombreux céphalopodes sont des créatures sociales ; lorsqu'elles sont isolées de leur propre espèce, certaines espèces ont été observées se regroupant en bancs avec des poissons.
Certains céphalopodes sont capables de voler sur des distances allant jusqu'à *Todarodes pacificus* , a été observée déployant ses tentacules en éventail, séparés par un film de mucus , tandis qu'une autre, *Sepioteuthis sepioidea* , les dispose en cercle.
Les sens
Les céphalopodes possèdent une vision avancée, peuvent détecter la gravité grâce à leurs statocystes et disposent de divers organes sensoriels chimiques. Les pieuvres utilisent leurs bras pour explorer leur environnement et peuvent s'en servir pour la perception de la profondeur.
Vision
L'œil primitif du nautile fonctionne de manière similaire à un appareil photo sténopé .La pupille en forme de W de la seiche se dilate lorsque les lumières s'éteignent.
La plupart des céphalopodes utilisent la vision pour détecter leurs prédateurs et leurs proies, et pour communiquer entre eux. Par conséquent, leur vision est très fine : des expériences d’entraînement ont montré que la pieuvre commune peut distinguer la luminosité, la taille, la forme et l’orientation horizontale ou verticale des objets. La morphologie de leurs yeux leur confère des performances similaires à celles des yeux de requins ; cependant, leur structure diffère, car les céphalopodes sont dépourvus de cornée et possèdent une rétine éversée. Les yeux des céphalopodes sont également sensibles au plan de polarisation de la lumière. Contrairement à de nombreux autres céphalopodes, les nautiles n’ont pas une bonne vision ; la structure de leur œil est très développée, mais ils sont dépourvus de cristallin . Ils possèdent un simple œil « à trou d’épingle » par lequel l’eau peut passer. On pense que cet animal utilise l’olfaction comme sens principal pour se nourrir , ainsi que pour localiser ou identifier des partenaires potentiels.
Une seiche dotée de pupilles en forme de W, ce qui pourrait l'aider à distinguer les couleurs.
Tous les poulpes et la plupart des céphalopodes sont considérés comme daltoniens . Les céphalopodes coléoïdes (poulpes, calmars, seiches) possèdent un seul type de photorécepteur et sont incapables de déterminer les couleurs en comparant l'intensité des photons détectés sur plusieurs canaux spectraux. Pour se camoufler , ils utilisent leurs chromatophores afin de modifier leur luminosité et leur motif en fonction de l'arrière-plan, mais leur capacité à imiter la couleur spécifique d'un arrière-plan pourrait provenir de cellules telles que les iridophores et les leucophores, qui réfléchissent la lumière ambiante . Ils produisent également des pigments visuels dans tout leur corps et pourraient percevoir directement les niveaux de lumière . Des indices de vision des couleurs ont été observés chez le calmar scintillant ( Watasenia scintillans ). La vision des couleurs est assurée par trois photorécepteurs , basés sur la même opsine , mais utilisant des molécules rétiniennes distinctes comme chromophores : A1 (rétinal), A3 (3-déshydrorétinal) et A4 (4-hydroxyrétinal). Le photorécepteur A1 est plus sensible au vert-bleu (484 nm), le photorécepteur A2 au bleu-vert (500 nm) et le photorécepteur A4 à la lumière bleue (470 nm).
En 2015, un nouveau mécanisme de discrimination spectrale chez les céphalopodes a été décrit. Ce mécanisme repose sur l'exploitation de l'aberration chromatique (dépendance de la distance focale à la longueur d'onde). La modélisation numérique montre que l'aberration chromatique peut fournir des informations chromatiques utiles grâce à la dépendance de l'acuité visuelle à l'accommodation. La forme inhabituelle de la fente oculaire hors axe et de la pupille annulaire chez les céphalopodes renforce cette capacité en agissant comme des prismes qui diffusent la lumière blanche dans toutes les directions.
Photoréception
En 2015, des données moléculaires ont été publiées indiquant que les chromatophores des céphalopodes sont photosensibles ; la RT-PCR a révélé la présence de transcrits codant pour la rhodopsine et calmar côtier à longues nageoires ( Doryteuthis pealeii ), ainsi que chez la seiche commune ( Sepia officinalis ) et la seiche à larges massues ( Sepia latimanus ). Les auteurs affirment qu’il s’agit de la première preuve que les tissus dermiques des céphalopodes possèdent la combinaison de molécules nécessaire pour répondre à la lumière.
Audience
Il a été démontré que certaines pieuvres et certains calmars détectent le son à l'aide de leurs statocystes , mais, en général, les céphalopodes sont sourds.
Utilisation de la lumière
Cette seiche à larges bandes ( Sepia latimanus ) peut passer de teintes camouflage beiges et brunes (en haut) à jaune avec des reflets foncés (en bas) en moins d'une seconde.d'iridophores, de leucophores , de chromatophores et, chez certaines espèces, de photophores . Les chromatophores sont des cellules pigmentaires colorées qui se dilatent et se contractent pour produire des couleurs et des motifs, qu'ils peuvent utiliser de manières étonnantes. Outre le camouflage qu'ils offrent, certains céphalopodes sont bioluminescents : ils émettent de la lumière vers le bas pour dissimuler leur ombre et la rendre invisible aux prédateurs qui pourraient se cacher en dessous. Cette bioluminescence est produite par des bactéries symbiotiques ; le céphalopode hôte est capable de détecter la lumière émise par ces organismes. La bioluminescence peut également servir à attirer les proies, et certaines espèces utilisent des parades colorées pour impressionner leurs partenaires, surprendre les prédateurs, ou même communiquer entre elles.
Coloration
la signalisation ( au sein de l'espèce ou pour avertir ) ou le camouflage actif , grâce à l'expansion ou la contraction de leurs chromatophores . Bien que les changements de couleur semblent dépendre principalement de la vision, il existe des preuves que les cellules cutanées, et plus particulièrement les chromatophores , peuvent détecter la lumière et s'adapter aux conditions d'éclairage indépendamment des yeux . La pieuvre modifie la couleur et la texture de sa peau au cours de ses cycles de sommeil calme et actif
Les céphalopodes peuvent utiliser leurs chromatophores comme des muscles, ce qui explique leur capacité à modifier rapidement la couleur de leur peau. La coloration est généralement plus intense chez les espèces côtières que chez celles vivant en haute mer, dont les fonctions se limitent souvent à un camouflage disruptif . Ces chromatophores sont présents dans tout le corps de la pieuvre ; cependant, ils sont contrôlés par la même partie du cerveau qui contrôle l’élongation lors de la propulsion par jet pour réduire la résistance. Ainsi, les pieuvres propulsées par jet peuvent devenir pâles car leur cerveau est incapable de contrôler simultanément l’élongation et les chromatophores. La plupart des pieuvres imitent certaines structures de leur champ de vision plutôt que d’adopter une couleur composite de leur environnement.
Des traces de coloration originelle ont été détectées chez des céphalopodes fossiles datant du Silurien ; ces individus orthocones présentaient des rayures concentriques, probablement utilisées comme camouflage. Les céphalopodes du Dévonien arborent des motifs de coloration plus complexes, dont la fonction reste inconnue.
Chromatophores
Les coléoïdes, une sous-classe de céphalopodes sans coquille (calmars, seiches et pieuvres), possèdent des cellules pigmentées complexes appelées chromatophores, capables de produire des motifs colorés changeants. Ces cellules stockent le pigment dans un sac élastique qui produit la couleur perçue. Les coléoïdes peuvent modifier la forme de ce sac, appelé sacculus cytoélastique, ce qui induit des changements de translucidité et d'opacité de la cellule. En modifiant rapidement la couleur de plusieurs chromatophores de différentes teintes, les céphalopodes peuvent changer la couleur de leur peau à une vitesse étonnante, une adaptation particulièrement remarquable chez un organisme qui voit en noir et blanc. On sait que les chromatophores ne contiennent que trois pigments, rouge, jaune et brun, ce qui ne permet pas de reproduire tout le spectre des couleurs. Cependant, les céphalopodes possèdent également des cellules appelées iridophores, de fines cellules protéiques stratifiées qui réfléchissent la lumière de manière à produire des couleurs que les chromatophores ne peuvent pas reproduire. Le mécanisme de contrôle des iridophores est inconnu, mais les chromatophores sont contrôlés par des voies neuronales, permettant ainsi au céphalopode de coordonner des présentations élaborées. Ensemble, chromatophores et iridophores sont capables de produire une large gamme de couleurs et de motifs.
Valeur adaptative
Les céphalopodes utilisent la capacité de leurs chromatophores à changer de couleur pour se camoufler. Les chromatophores permettent aux céphalopodes de se fondre dans des environnements très variés, des récifs coralliens aux fonds marins sableux. Le changement de couleur des chromatophores agit de concert avec les papilles, un tissu épithélial qui se développe et se déforme sous l'effet de mouvements hydrostatiques, modifiant ainsi la texture de la peau. Les chromatophores sont capables de deux types de camouflage : le mimétisme et l'homochromie. Le mimétisme consiste pour un organisme à modifier son apparence pour ressembler à un autre. On a observé que le calmar Sepioteuthis sepioidea change d'apparence pour se faire passer pour le poisson-perroquet herbivore inoffensif et approcher des proies sans méfiance. La pieuvre Thaumoctopus mimicus est connue pour imiter plusieurs organismes venimeux avec lesquels elle cohabite afin de dissuader les prédateurs. Dans le cas de l'homochromie, un céphalopode modifie son apparence pour se fondre dans son environnement, se cachant ainsi de ses prédateurs ou se dissimulant à ses proies. La capacité à la fois d'imiter d'autres organismes et de reproduire l'apparence de leur environnement est remarquable étant donné que la vision des céphalopodes est monochrome.
Les céphalopodes utilisent également le contrôle précis de la coloration et des motifs de leur corps pour réaliser des signaux complexes, tant au sein de leur propre espèce qu'entre individus de la même espèce. La coloration, combinée à la locomotion et à la texture, permet d'envoyer des signaux aux autres organismes. Au sein d'une même espèce, il peut s'agir d'un signal d'avertissement pour les prédateurs potentiels. Par exemple, lorsqu'une pieuvre (Callistoctopus macropus) se sent menacée, elle prend une couleur brun-rouge vif parsemée de points blancs, créant un contraste élevé pour surprendre les prédateurs. Au sein d'une même espèce, le changement de couleur est utilisé pour les parades nuptiales et la communication sociale. Les seiches, par exemple, présentent des parades nuptiales élaborées entre mâles et femelles. Il existe également une signalisation entre mâles lors de la compétition pour les partenaires, tous ces phénomènes étant le fruit de la coloration des chromatophores.
Origine
Deux hypothèses expliquent l'évolution du changement de couleur chez les céphalopodes. Selon la première, cette capacité aurait évolué pour des raisons sociales, sexuelles et de communication. Selon la seconde, elle serait apparue initialement sous l'effet de pressions sélectives favorisant l'évitement des prédateurs et la chasse furtive.
Pour que le changement de couleur soit le résultat de la sélection sociale, l'environnement des ancêtres des céphalopodes aurait dû répondre à plusieurs critères. Premièrement, il aurait fallu un rituel d'accouplement impliquant une signalisation. Deuxièmement, une forte sélection sexuelle aurait dû être observée. Troisièmement, l'ancêtre aurait dû communiquer par des signaux sexuels visibles par un congénère. Pour que le changement de couleur soit le résultat de la sélection naturelle, d'autres paramètres auraient dû être réunis. Il aurait fallu une certaine diversité phénotypique dans les motifs corporels au sein de la population. L'espèce aurait également dû cohabiter avec des prédateurs qui utilisent la vision pour identifier leurs proies. Ces prédateurs auraient dû posséder une grande sensibilité visuelle, détectant non seulement le mouvement et le contraste, mais aussi les couleurs. Les habitats qu'ils occupent auraient également dû présenter une diversité de décors. Des expériences menées sur des caméléons nains pour tester ces hypothèses ont montré que les taxons de caméléons dotés d'une plus grande capacité de changement de couleur présentaient des signaux sociaux plus visibles, mais ne provenaient pas d'habitats visuellement plus diversifiés. Ceci suggère que la capacité de changement de couleur a probablement évolué pour faciliter la signalisation sociale, le camouflage étant un effet secondaire utile. Le camouflage étant utilisé à de multiples fins adaptatives chez les céphalopodes, le changement de couleur pourrait avoir évolué pour un usage initial, l'autre s'étant développé ultérieurement, ou bien il pourrait avoir évolué pour réguler les compromis entre les deux.
Évolution convergente
Le changement de couleur est fréquent chez les ectothermes, notamment les anolis, les grenouilles, les mollusques, de nombreux poissons, les insectes et les araignées. Le mécanisme sous-jacent à ce changement de couleur peut être morphologique ou physiologique. Le changement morphologique résulte d'une modification de la densité des cellules pigmentées et tend à évoluer sur de longues périodes. Le changement physiologique, tel qu'observé chez les céphalopodes, résulte généralement du déplacement des pigments au sein du chromatophore, modifiant ainsi leur localisation cellulaire. Ce changement physiologique se produit généralement sur des échelles de temps beaucoup plus courtes que le changement morphologique. Les céphalopodes présentent une forme rare de changement de couleur physiologique qui utilise le contrôle neuronal des muscles pour modifier la morphologie de leurs chromatophores. Ce contrôle neuronal des chromatophores a évolué de manière convergente chez les céphalopodes et les poissons téléostéens.
Encre
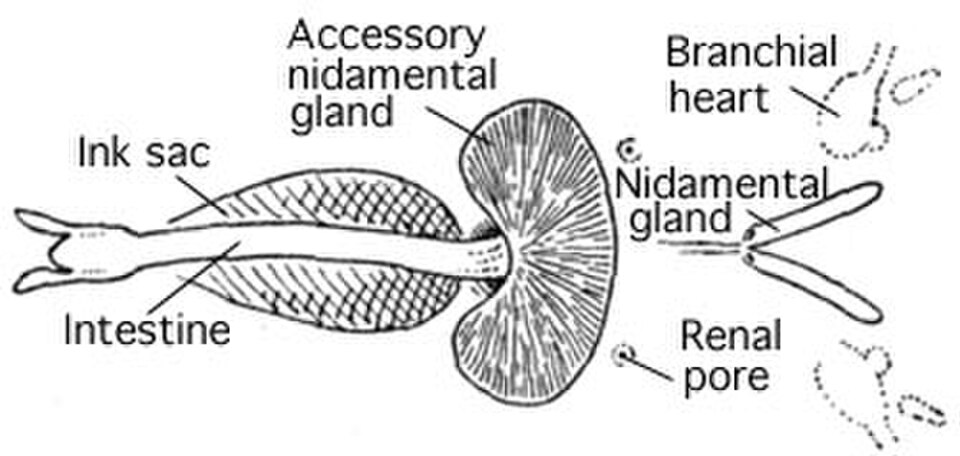
Nautilidae et des espèces de pieuvres appartenant au sous-ordre des Cirrina [ les céphalopodes connus possèdent une poche à encre, qui leur permet d'expulser un nuage d'encre foncée pour tromper leurs prédateurs . Cette poche est un sac musculaire issu d'une extension de l'intestin postérieur. Située sous l'intestin, elle s'ouvre dans l'anus, où son contenu – de la mélanine presque pure – est éjecté. Sa proximité avec la base de l'anus permet à l'encre d'être dispersée par l'eau expulsée lors de la propulsion par jet . Le nuage de mélanine éjecté se mélange généralement, lors de son expulsion, à du mucus produit ailleurs dans le manteau, formant ainsi un épais nuage qui altère la vision (et possiblement les sens chimiosensoriels) du prédateur, à la manière d'un écran de fumée . Cependant, un comportement plus sophistiqué a été observé : le céphalopode libère un nuage de mucus plus dense, ressemblant approximativement au céphalopode qui l’a libéré (ce leurre est appelé pseudomorphe ). Cette stratégie conduit souvent le prédateur à attaquer le pseudomorphe plutôt que sa proie qui s’éloigne rapidement. Pour plus d’informations, voir Comportements d’encrage .
La poche à encre des céphalopodes a donné lieu à un nom commun de « poisson à encre », anciennement poisson à l'encre.
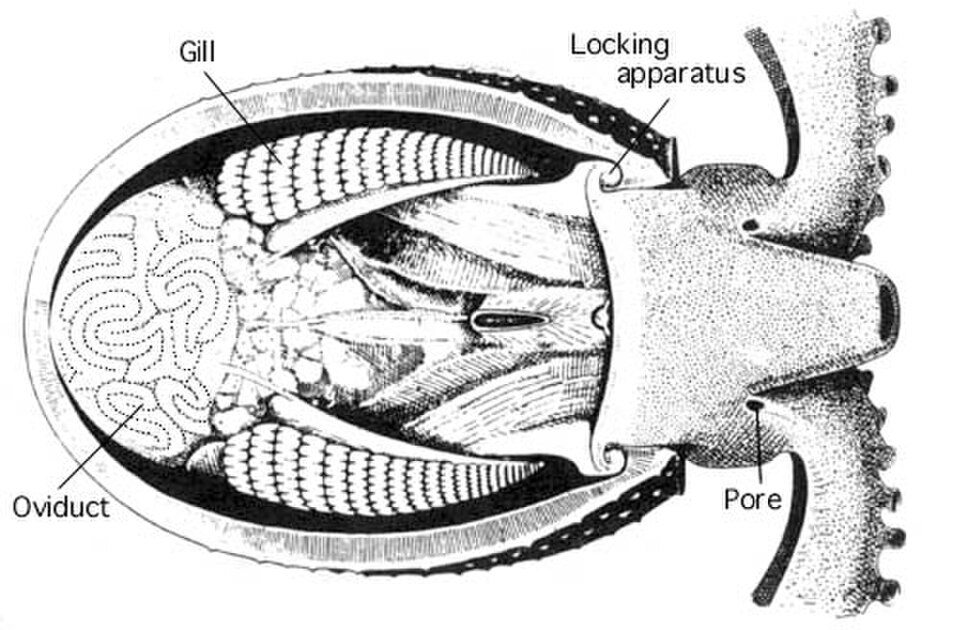
Les céphalopodes sont les seuls mollusques à posséder un système circulatoire clos. Les coléoïdes ont deux cœurs branchiaux qui font circuler le sang à travers les capillaires des branchies . Un seul cœur systémique pompe ensuite le sang oxygéné vers le reste du corps.
Comme la plupart des mollusques, les céphalopodes utilisent l'hémocyanine , une protéine contenant du cuivre, plutôt que l'hémoglobine , pour transporter l'oxygène. De ce fait, leur sang est incolore lorsqu'il est désoxygéné et devient bleu lorsqu'il se lie à l'oxygène. Dans les milieux riches en oxygène et en eau acide, l'hémoglobine est plus efficace, mais dans les milieux pauvres en oxygène et à basses températures, l'hémocyanine est prépondérante. La molécule d'hémocyanine est beaucoup plus grosse que la molécule d'hémoglobine, ce qui lui permet de se lier à 96 molécules d' O₂ ou de Les pieuvres nagent la tête la première, les bras traînant derrière.
Bien que la plupart des céphalopodes puissent se déplacer par propulsion à réaction , ce mode de déplacement est très énergivore comparé à la propulsion caudale utilisée par les poissons. L'efficacité d'un hydrojet entraîné par une hélice (c'est-à-dire l'efficacité de Froude ) est supérieure à celle d'une fusée . L'efficacité relative de la propulsion à réaction diminue encore avec la taille de l'animal ; les paralarves sont bien plus efficaces que les juvéniles et les adultes. Depuis l' ère paléozoïque , la compétition avec les poissons ayant engendré un environnement où un déplacement efficace était crucial pour la survie, la propulsion à réaction est devenue secondaire, les nageoires et les tentacules servant alors à maintenir une vitesse constante. Bien que la propulsion à réaction ne soit jamais le seul mode de locomotion, le mouvement saccadé qu'elle permet reste utile pour atteindre des vitesses élevées, notamment pour capturer des proies ou échapper aux prédateurs . En effet, cela fait des céphalopodes les invertébrés marins les plus rapides, , et ils peuvent accélérer plus vite que la plupart des poissons. Le jet est complété par le mouvement des nageoires ; chez le calmar, les nageoires battent à chaque jet, amplifiant la poussée ; elles sont ensuite déployées entre les jets (probablement pour éviter de couler). L’eau oxygénée est aspirée dans la cavité palléale jusqu’aux branchies et, par contraction musculaire de cette cavité, l’eau usée est expulsée par l’ hyponome , formé par un repli du manteau. La différence de taille entre les extrémités postérieure et antérieure de cet organe détermine la vitesse du jet que l’organisme peut produire. La vitesse de l’organisme peut être prédite avec précision pour une masse et une morphologie animales données. Le mouvement des céphalopodes est généralement vers l’arrière, l’eau étant expulsée vers l’avant par l’hyponome, mais la direction peut être contrôlée dans une certaine mesure en l’orientant dans différentes directions. Certains céphalopodes accompagnent cette expulsion d'eau d'un bruit sec ressemblant à un coup de feu, que l'on pense destiné à effrayer les prédateurs potentiels.
Les céphalopodes utilisent une méthode de propulsion similaire malgré leur taille croissante (qui modifie la dynamique de l'eau dans laquelle ils évoluent). Ainsi, leurs paralarves n'utilisent pas beaucoup leurs nageoires (moins efficaces à faible nombre de Reynolds ) et se propulsent principalement vers le haut grâce à leurs jets d'eau, tandis que les grands céphalopodes adultes ont tendance à nager moins efficacement et à s'appuyer davantage sur leurs nageoires.
On pense que les premiers céphalopodes produisaient des jets en rétractant leur corps dans leur coquille, comme le fait le nautile aujourd'hui. Le nautile est également capable de créer un jet par ondulation de son conduit ; ce flux d'eau plus lent est mieux adapté à l'extraction d'oxygène. À l'arrêt, le nautile ne peut extraire que 20 % de l'oxygène de l'eau. La vitesse du jet chez le nautile est beaucoup plus faible que chez les coléoïdes , mais sa production requiert moins de musculature et d'énergie. La propulsion par jet chez les céphalopodes est principalement contrôlée par le diamètre maximal de l'orifice du conduit (ou peut-être par son diamètre moyen) et par le diamètre de la cavité palléale. Les variations de taille de l'orifice sont surtout utilisées aux vitesses intermédiaires. La vitesse absolue atteinte est limitée par la nécessité pour le céphalopode d'inhaler de l'eau pour l'expulser ; cette ingestion limite la vitesse maximale à huit longueurs de corps par seconde, une vitesse que la plupart des céphalopodes peuvent atteindre après deux coups d'entonnoir. L'eau remplit la cavité en entrant non seulement par les orifices, mais aussi par l'entonnoir. Le calmar peut expulser jusqu'à 94 % du fluide contenu dans sa cavité en une seule poussée. Pour s'adapter aux variations rapides d'ingestion et d'expulsion d'eau, les orifices sont très flexibles et leur taille peut varier d'un facteur 20 ; le rayon de l'entonnoir, en revanche, ne varie que d'un facteur d'environ 1,5.
Certaines espèces de pieuvres sont également capables de marcher sur le fond marin. Les calmars et les seiches peuvent se déplacer sur de courtes distances dans toutes les directions en ondulant un repli musculaire situé autour de leur manteau.
Bien que la plupart des céphalopodes flottent (c’est-à-dire qu’ils ont une flottabilité neutre ou quasi neutre ; en fait, la plupart des céphalopodes sont environ 2 à 3 % plus denses que l’eau de mer ), ils y parviennent de différentes manières. Certains, comme le nautile , permettent aux gaz de diffuser dans l’espace entre le manteau et la coquille ; d’autres laissent l’eau plus pure suinter de leurs reins, chassant ainsi l’eau salée plus dense de la cavité corporelle ; d’autres encore, comme certains poissons, accumulent des huiles dans le foie ; et certaines pieuvres ont un corps gélatineux où les ions chlorure, plus légers, remplacent les ions sulfate dans leur composition chimique.
Les calmars sont les principaux céphalopodes affectés par la flottabilité négative. Cette flottabilité négative signifie que certains calmars, notamment ceux vivant en eaux peu profondes, doivent réguler activement leur position verticale. Ils doivent donc dépenser de l'énergie, souvent par des jets d'eau ou des ondulations, pour maintenir une profondeur constante. De ce fait, le coût énergétique du transport est assez élevé pour de nombreux calmars. Cela dit, les calmars et autres céphalopodes vivant en eaux profondes ont généralement une flottabilité plus neutre, ce qui les dispense de réguler leur profondeur et améliore leur efficacité locomotrice.
Le Macrotritopus defilippi , ou pieuvre des sables, a été observé imitant la coloration et les mouvements de nage du flet des sables Bothus lunatus pour échapper aux prédateurs. Les pieuvres étaient capables d'aplatir leur corps et de replier leurs bras pour ressembler aux flets et se déplacer avec la même vitesse et les mêmes mouvements.
Les femelles de deux espèces, Ocythoe tuberculata et Haliphron atlanticus , ont développé une véritable vessie natatoire .
locomotion du poulpe contre celle du calmar
Deux catégories de céphalopodes, les pieuvres et les calmars, présentent des modes de déplacement très différents malgré leur appartenance à la même classe. Les pieuvres ne sont généralement pas considérées comme des nageuses actives ; on les observe souvent fouiller le fond marin plutôt que de parcourir de longues distances. Les calmars, en revanche, peuvent parcourir d'immenses distances, certains parcourant jusqu'à 2 000 km en 2,5 mois à une vitesse moyenne de 0,9 longueur de corps par seconde. Cette différence de type et d'efficacité de déplacement s'explique principalement par l'anatomie.
Les pieuvres et les calmars possèdent tous deux un manteau (mentionné précédemment) qui assure la respiration et la locomotion par propulsion. La composition de ce manteau diffère cependant entre les deux familles. Chez les pieuvres, le manteau est constitué de trois types de muscles : longitudinaux, radiaux et circulaires. Les muscles longitudinaux, parallèles à l’axe du corps, permettent de maintenir la longueur du manteau constante lors de la propulsion. De ce fait, la pieuvre doit les contracter activement pendant la propulsion afin de maintenir cette longueur. Les muscles radiaux, perpendiculaires aux muscles longitudinaux, servent à moduler l’épaisseur du manteau. Enfin, les muscles circulaires, principaux activateurs de la propulsion, sont des bandes musculaires qui entourent le manteau et permettent de dilater ou de contracter la cavité. Ces trois types de muscles agissent de concert pour produire un jet d’eau servant de mécanisme de propulsion.
Les calmars ne possèdent pas de muscles longitudinaux comme les pieuvres. Ils sont recouverts d'une tunique . Cette tunique, composée de couches de collagène, enveloppe le manteau par le haut et par le bas. Grâce à sa structure en collagène et non en muscle, la tunique est une structure rigide et beaucoup plus résistante que les muscles. Cette rigidité confère aux calmars des avantages pour la propulsion par jet. Elle leur permet notamment de maintenir le manteau de taille constante sans avoir à contracter leurs muscles. De plus, la tunique ne représente que 1 % de l'épaisseur du manteau du calmar, tandis que les fibres musculaires longitudinales peuvent représenter jusqu'à 20 % de cette épaisseur chez la pieuvre . Enfin, la rigidité de la tunique permet aux muscles radiaux du calmar de se contracter avec plus de force.
Le manteau n'est pas le seul endroit où l'on trouve du collagène chez les calmars. Des fibres de collagène sont présentes dans l'ensemble des autres fibres musculaires du manteau. Ces fibres de collagène agissent comme des élastiques et sont parfois appelées « ressorts de collagène » . Comme leur nom l'indique, ces fibres fonctionnent comme des ressorts. Lorsque les muscles radiaux et circulaires du manteau se contractent, ils atteignent un point où la contraction n'est plus efficace pour la propulsion de l'animal. Dans ce cas, l'excédent de contraction est stocké dans le collagène, qui amorce ou contribue alors efficacement à l'expansion du manteau en fin de course. Lors de certains tests, il a été démontré que le collagène pouvait commencer à augmenter la pression du manteau jusqu'à 50 ms avant le début de l'activité musculaire
Ces différences anatomiques entre les calmars et les pieuvres peuvent aider à expliquer pourquoi les calmars peuvent être trouvés en train de nager de manière comparable aux poissons alors que les pieuvres s'appuient généralement sur d'autres formes de locomotion sur le fond marin telles que la marche bipède, le reptation et la nage sans jet.
Coquille
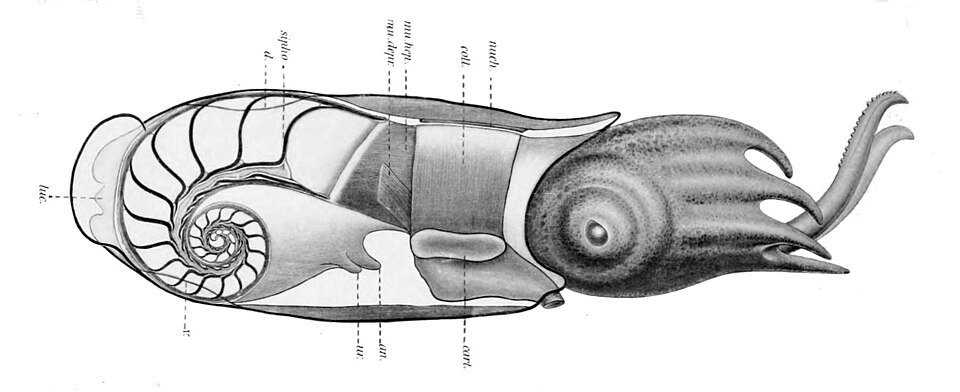
Coupe transversale de Spirula spirula , montrant la position de la coquille à l'intérieur du manteau
Les nautiles sont les seuls céphalopodes actuels à posséder une véritable coquille externe. Cependant, toutes les coquilles de mollusques sont formées à partir de l' ectoderme (couche externe de l'embryon) ; chez les seiches ( Sepia spp.), par exemple, une invagination de l'ectoderme se forme durant la période embryonnaire, aboutissant à une coquille ( l'os de seiche ) interne à l'adulte. Il en va de même pour le gladius chitineux des calmars et des pieuvres. Les pieuvres cirrées possèdent des supports cartilagineux en forme d'arche pour leurs nageoires , parfois appelés « vestiges de coquille » ou « gladius ». Les Incirrina possèdent soit une paire de stylets en forme de bâtonnet , soit aucun vestige de coquille interne, et certains calmars sont également dépourvus de gladius. Les coléoïdes à coquille ne forment ni un clade ni même un groupe paraphylétique. La coquille de Spirula se forme initialement à partir d'une structure organique, puis se minéralise très rapidement. Les coquilles « perdues » le sont probablement par résorption de leur composant carbonaté.
Les femelles du genre de pieuvres Argonauta sécrètent une capsule d'œufs spécialisée, aussi fine que du papier, dans laquelle elles résident, et celle-ci est communément considérée comme une « coquille », bien qu'elle ne soit pas attachée au corps de l'animal et ait une origine évolutive distincte.
Les ammonites , le plus grand groupe de céphalopodes à coquille , sont éteints, mais leurs coquilles sont très fréquentes à l' état fossile . Les ammonites ont prospéré durant les ères paléozoïque et mésozoïque. Leurs coquilles caractéristiques, en forme de spirale, se retrouvent dans les roches sédimentaires du monde entier et, par conséquent, dans de nombreuses créations humaines.
Le dépôt de carbonate, conduisant à une coquille minéralisée, semble lié à l'acidité de la matrice organique de la coquille (voir Coquille de mollusque ) ; les céphalopodes à coquille possèdent une matrice acide, tandis que le gladius des calmars possède une matrice basique. La structure de base de la paroi externe des céphalopodes est la suivante : une couche prismatique externe (sphérolitique), une couche laminaire (nacrée) et une couche prismatique interne. L'épaisseur de chaque couche varie selon les taxons. Chez les céphalopodes modernes, le carbonate de calcium est de l'aragonite. Comme pour les autres coquilles de mollusques ou les squelettes coralliens, les plus petites unités visibles sont des granules arrondis irréguliers.
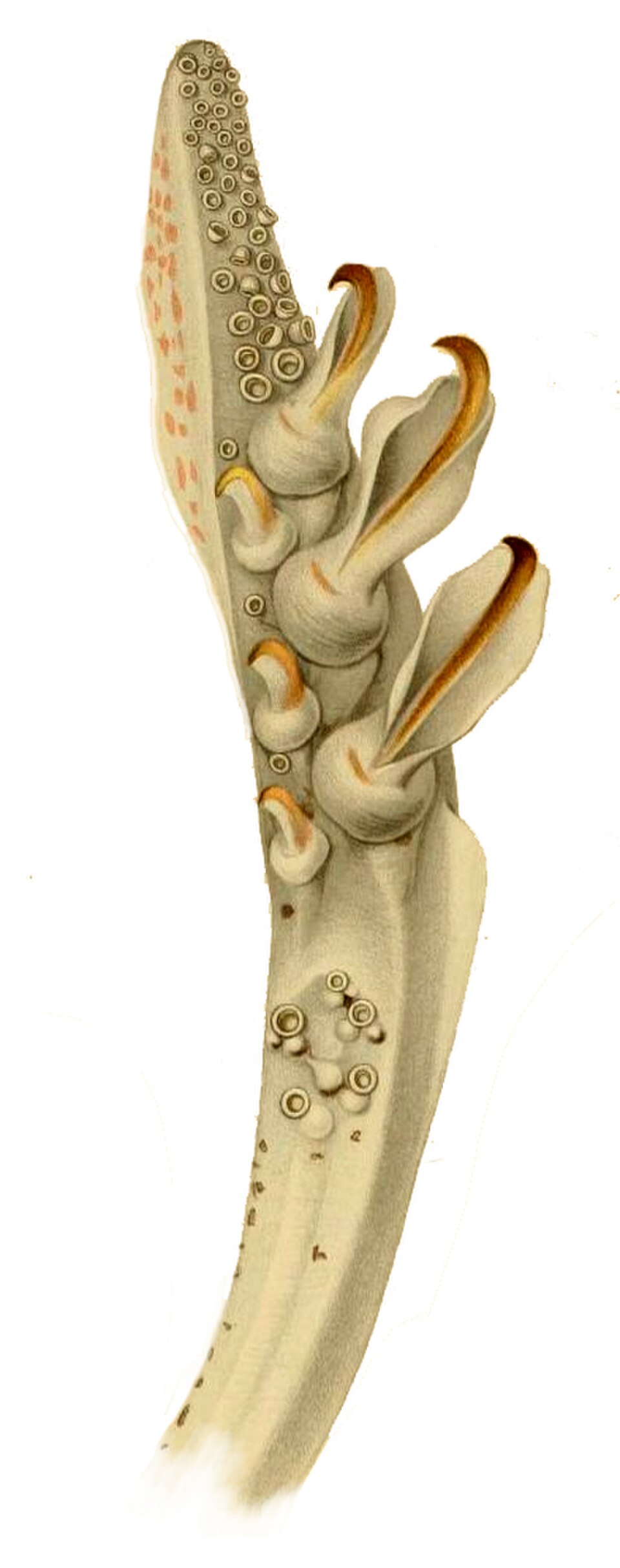
À gauche : Un calmar géant découvert à Logy Bay , à Terre-Neuve , en 1873. Les deux longs tentacules nourriciers sont visibles à l'extrême gauche et à droite. À droite : Détail du club tentaculaire d' Abraliopsis morisi
appendices céphaliques
les coléoïdes, on en compte huit ou dix paires. Les décapodes, tels que les seiches et les calmars, en possèdent cinq. Les deux paires les plus longues, appelées « tentacules », participent activement à la capture des proies ; elles peuvent s'allonger rapidement (en seulement 15 millisecondes ). Chez le calmar géant , elles peuvent atteindre une longueur de 8 mètres. Elles peuvent se terminer par une massue élargie recouverte de ventouses. Les quatre paires les plus courtes sont appelées bras et servent à maintenir et à manipuler l'organisme capturé. elles aussi sont munies de ventouses, du côté le plus proche de la bouche ; celles-ci permettent de retenir la proie. Les octopodes n'ont que quatre paires de bras recouverts de ventouses, comme leur nom l'indique, bien que des anomalies de développement puissent modifier le nombre de bras exprimés.
Le tentacule est constitué d'un épais cordon nerveux central (dont l'épaisseur est nécessaire pour permettre le contrôle indépendant de chaque ventouse) entouré de muscles circulaires et radiaux. Le volume du tentacule restant constant, la contraction des muscles circulaires diminue le rayon et permet un allongement rapide. Typiquement, un allongement de 70 % est obtenu par une réduction de la largeur de 23 %. Les bras plus courts sont dépourvus de cette capacité.
La taille du tentacule est liée à la taille de la cavité buccale ; des tentacules plus grands et plus forts peuvent retenir la proie pendant qu’ils en prélèvent de petites bouchées ; avec des tentacules plus nombreux et plus petits, la proie est avalée entière, la cavité buccale doit donc être plus grande.
Les nautiles à coquille externe ( Nautilus et Allonautilus ) possèdent environ 90 appendices en forme de doigts, appelés tentacules , qui sont dépourvus de ventouses mais collants et partiellement rétractables.
Alimentation
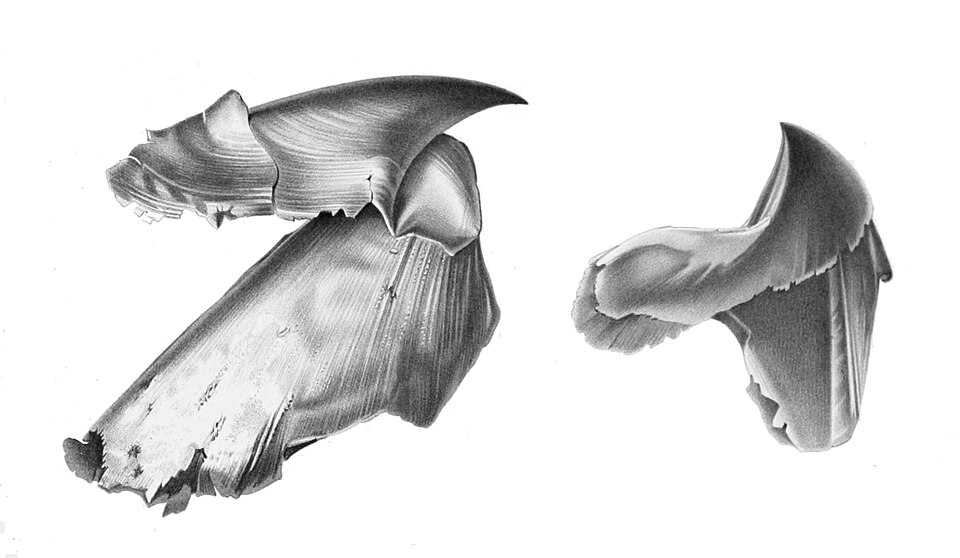
Le bec en deux parties du calmar géant , Architeuthis sp.
Tous les céphalopodes vivants possèdent un bec en deux parties ; la plupart ont une radula , bien que celle-ci soit réduite chez la plupart des pieuvres et totalement absente chez les spirules . Ils se nourrissent en capturant leurs proies avec leurs tentacules, en les attirant dans leur bouche et en les mordant. Ils possèdent un mélange de sucs digestifs toxiques, dont certains sont produits par des algues symbiotiques, qu’ils projettent de leurs glandes salivaires sur leurs proies capturées. Ces sucs séparent la chair de leurs proies de l’os ou de la coquille. La glande salivaire possède une petite dent à son extrémité qui peut être enfoncée dans un organisme pour le digérer de l’intérieur.
La glande digestive elle-même est relativement courte. Elle comprend quatre parties : le jabot, l’estomac et le cæcum, avant que les aliments n’atteignent l’intestin. La majeure partie de la digestion, ainsi que l’absorption des nutriments, se déroule dans la glande digestive, parfois appelée foie. Les nutriments et les déchets sont échangés entre l’intestin et la glande digestive grâce à deux connexions reliant cette dernière à la jonction de l’estomac et du cæcum. Les cellules de la glande digestive libèrent directement des substances chimiques pigmentées dans la lumière intestinale. Ces substances sont ensuite liées au mucus et évacuées par l’anus sous forme de longs filaments sombres, expulsés avec l’aide de l’eau expirée. Les céphalopodes ont tendance à concentrer les métaux lourds ingérés dans leurs tissus. Cependant, les tentacules des pieuvres utilisent une famille de récepteurs chimioactiles (RC) spécifiques aux céphalopodes pour leur système de « goût par le toucher ».
La radula des céphalopodes est constituée de plusieurs rangées symétriques comportant jusqu'à neuf dents – treize chez les espèces fossiles. Cet organe est réduit, voire vestigial, chez certaines espèces de pieuvres et est absent chez Spirula . Les dents peuvent être homodontes (c'est-à-dire de forme similaire sur toute la rangée), hétérodontes (c'est-à-dire de forme différente) ou cténodontes (en forme de peigne). Leur hauteur, leur largeur et le nombre de cuspides varient selon les espèces. La disposition des dents se répète, mais chaque rangée peut différer de la précédente ; chez la pieuvre, par exemple, la séquence se répète tous les cinq rangs.
Les radulas de céphalopodes sont connues grâce à des gisements fossilifères datant de l'Ordovicien. Elles sont généralement conservées dans la loge du céphalopode, souvent associées aux mandibules ; mais ce n'est pas toujours le cas ; de nombreuses radulas sont conservées dans divers environnements du Mason Creek. Les radulas sont généralement difficiles à détecter, même lorsqu'elles sont fossilisées, car la roche doit s'altérer et se fissurer de manière très précise pour les exposer ; par exemple, des radulas n'ont été trouvées que dans neuf des 43 genres d'ammonites, et elles sont encore plus rares chez les formes non ammonites : seules trois espèces pré-mésozoïques en possèdent une.
Système excréteur
La plupart des céphalopodes possèdent une seule paire de grands néphridies . Les déchets azotés filtrés sont produits dans la cavité péricardique des cœurs branchiaux , chacun étant relié à une néphridie par un canal étroit. Ce canal achemine les excréments vers un sac rénal vésiculaire et réabsorbe également l'excès d'eau du filtrat. Plusieurs excroissances de la veine cave latérale se projettent dans le sac rénal, se gonflant et se dégonflant continuellement au rythme des battements des cœurs branchiaux. Ce mécanisme permet de pomper les déchets sécrétés dans les sacs, d'où ils sont libérés dans la cavité palléale par un pore.
Le nautile , fait inhabituel, possède quatre néphridies, dont aucune n'est reliée aux cavités péricardiques.
L'incorporation d' ammoniac est importante pour la formation de la coquille chez les mollusques terrestres et d'autres lignées non mollusques. Les protéines (c'est-à-dire la chair) constituant une part importante du régime alimentaire des céphalopodes, de grandes quantités d' ions ammonium sont produites comme déchets. Les branchies sont les principaux organes impliqués dans l'élimination de cet excès d'ammonium. Le taux d'élimination est le plus faible chez les céphalopodes à coquille comme le nautile et la seiche , car ils utilisent l'azote pour remplir leur coquille de gaz et ainsi augmenter leur flottabilité. D'autres céphalopodes utilisent l'ammonium de manière similaire, en stockant les ions (sous forme de chlorure d'ammonium ) afin de réduire leur densité globale et d'augmenter leur flottabilité.
Reproduction et cycle de vie
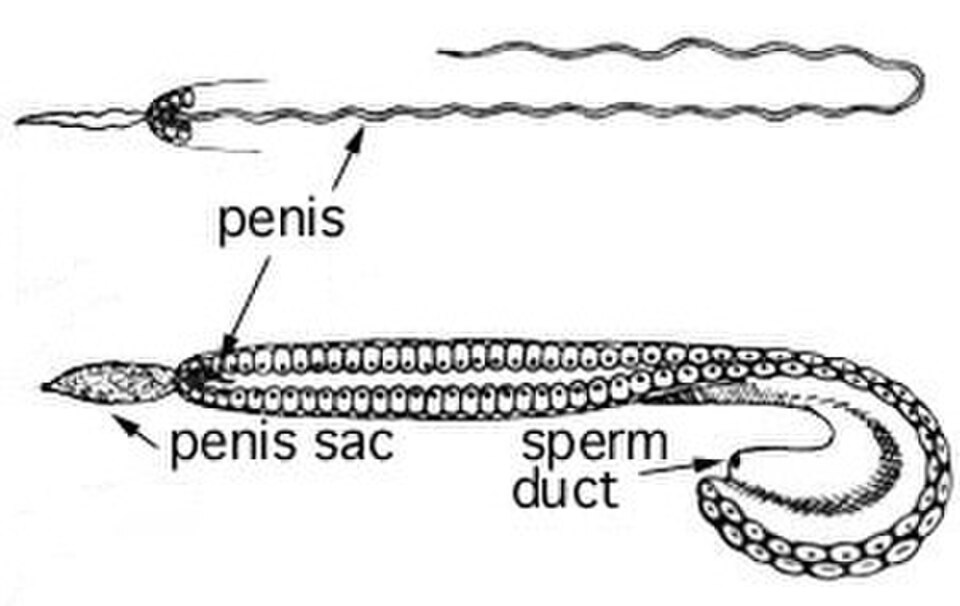
Femelle d'Argonauta argo avec oothèque et œufsDétail de l' hectocotyle d' Ocythoe tuberculataUn spécimen mâle disséqué d' Onykia ingens , montrant un pénis non érigé (la structure tubulaire blanche située sous la plupart des autres organes).Un spécimen de la même espèce présentant un allongement du pénis jusqu'à 67 cm de longueur
Les céphalopodes forment un groupe d'espèces diversifié, mais partagent des caractéristiques communes de leur cycle de vie, comme une croissance rapide et une courte durée de vie. Stearns (1992) a suggéré que, pour produire le plus grand nombre possible de descendants viables, la ponte dépend des facteurs environnementaux de l'organisme. La majorité des céphalopodes n'assurent pas de soins parentaux à leur progéniture, à l'exception, par exemple, de la pieuvre, ce qui contribue à augmenter le taux de survie de sa descendance. Les cycles de vie des espèces marines sont affectés par diverses conditions environnementales. Le développement d'un embryon de céphalopode peut être fortement influencé par la température, la saturation en oxygène, la pollution, l'intensité lumineuse et la salinité. Ces facteurs sont importants pour le rythme du développement embryonnaire et le succès de l'éclosion. La disponibilité alimentaire joue également un rôle important dans le cycle de reproduction des céphalopodes. Une limitation des ressources alimentaires influence le moment de la ponte, ainsi que le fonctionnement et la croissance des embryons. La période et la durée de la ponte varient selon les espèces marines ; elles sont corrélées à la température, bien que les céphalopodes vivant en eaux peu profondes se reproduisent pendant les mois froids afin que les alevins éclosent à des températures plus élevées. La reproduction peut durer de quelques jours à un mois.
maturité sexuelle
Les céphalopodes sexuellement matures et de taille adulte commencent à se reproduire. Après la transmission du matériel génétique à la génération suivante, la plupart des céphalopodes adultes meurent. La maturation sexuelle chez les céphalopodes mâles et femelles s'observe par l'hypertrophie des gonades et des glandes annexes. L'accouplement est un indicateur peu fiable de la maturation sexuelle chez les femelles ; celles-ci peuvent recevoir du sperme avant d'être pleinement matures et le stocker jusqu'à la fécondation des œufs. Les mâles sont plus agressifs lors des compétitions prénuptiales en présence de femelles immatures qu'avec une femelle mature. La plupart des céphalopodes mâles développent un hectocotyle, une extrémité de bras capable de transférer leurs spermatozoïdes dans la cavité palléale de la femelle. Cependant, toutes les espèces n'utilisent pas d'hectocotyle ; par exemple, le nautile adulte libère un spadice . Certains calmars mâles, principalement des espèces d'eaux profondes, ont développé un pénis plus long que leur propre corps ; il s'agit du pénis le plus long observé chez les animaux vivant en liberté. On suppose que ces mâles fixent simplement un spermatophore n'importe où sur le corps de la femelle. Le développement de photophores brachiaux, destinés à attirer les partenaires, est un indicateur de la maturité sexuelle des femelles.
Fertilisation
Les céphalopodes ne pratiquent pas la fécondation externe . Lors de la fécondation, les femelles utilisent le sperme fourni par le mâle . La fécondation interne n'est observée que chez les pieuvres. L'accouplement débute lorsque le mâle capture une femelle et l'entoure de son bras, soit par contact « cou contre cou », soit par contact bouche contre bouche, selon l'espèce. Le mâle initie ensuite la fécondation en contractant son manteau à plusieurs reprises pour libérer les spermatozoïdes. Les céphalopodes s'accouplent souvent plusieurs fois, ce qui incite les mâles à prolonger l'accouplement avec les femelles déjà accouplées, triplant presque le nombre de contractions du manteau. Pour assurer la fécondation des œufs, les femelles libèrent un peptide attractif pour les spermatozoïdes à travers les couches gélatineuses de l'œuf afin de guider ces derniers. Les femelles céphalopodes pondent leurs œufs par grappes. Chaque œuf est composé d'une enveloppe protectrice assurant la sécurité de l'embryon en développement lors de sa libération dans la colonne d'eau. Les stratégies de reproduction diffèrent selon les espèces de céphalopodes. Chez la pieuvre géante du Pacifique, les gros œufs sont pondus dans une tanière ; la ponte complète peut prendre plusieurs jours. Une fois les œufs libérés et généralement fixés à un substrat abrité, la femelle meurt généralement peu après, mais les pieuvres et quelques calmars continuent de veiller sur leurs œufs. D'autres espèces, comme le calmar volant japonais , pondent des masses d'œufs à flottabilité neutre qui flottent à l'interface entre des couches d'eau de densités légèrement différentes, ou bien la femelle nage en transportant les œufs. La plupart des espèces sont sémelpares (elles ne se reproduisent qu'une seule fois avant de mourir). Les seules exceptions connues sont le calmar vampire , la pieuvre rayée du Pacifique et le nautile , qui sont itéropares . Chez certaines espèces de céphalopodes, les pontes sont fixées au substrat par une substance adhésive mucilagineuse. Ces œufs sont gonflés de liquide périvitellin (LPV), un liquide hypertonique qui empêche l'éclosion prématurée. Les amas d'œufs fécondés ont une flottabilité neutre selon la profondeur de ponte, mais on peut aussi les trouver dans des substrats comme le sable, une matrice corallienne ou des algues. Comme ces espèces n'assurent pas de soins parentaux à leur progéniture, la femelle peut injecter de l'encre dans les capsules d'œufs afin de camoufler les embryons et les rendre invisibles aux prédateurs.
compétition entre mâlesLe choix du partenaire est observé chez les seiches, où les femelles préfèrent certains mâles à d'autres, bien que les caractéristiques de ces mâles préférés restent inconnues. Une hypothèse suggère que les femelles rejettent les mâles par des signaux olfactifs plutôt que visuels. Plusieurs espèces de céphalopodes sont polyandres : elles acceptent et stockent les spermatophores de plusieurs mâles, ce qui a été identifié par empreinte génétique. Les femelles ne sont plus réceptives aux tentatives d'accouplement lorsqu'elles portent leurs œufs dans leurs bras. Elles peuvent stocker le sperme à deux endroits : (1) la cavité buccale, où les mâles récemment accouplés déposent leurs spermatophores, et (2) les réceptacles internes de stockage du sperme, où sont stockés les spermatophores provenant de mâles précédents. Le stockage des spermatophores engendre une compétition spermatique : la femelle contrôle ainsi quel partenaire fécondera ses œufs. Afin de réduire cette compétition, les mâles développent des comportements agonistiques tels que la garde de la partenaire et le chassage des œufs. L' Hapalochlaena lunulata , ou la pieuvre à anneaux bleus, s'accouple facilement avec les mâles et les femelles.
dimorphisme sexuel
Chez divers organismes marins, on observe que les femelles sont plus grandes que les mâles chez certaines espèces étroitement apparentées. Dans certaines lignées, comme la pieuvre-couverture , les mâles deviennent structurellement plus petits, un phénomène appelé « nanisme ». Les mâles nains sont généralement présents en faibles densités. Le mâle de la pieuvre-couverture est un exemple de nanisme sexuel et évolutif ; les femelles atteignent une taille 10 000 à 40 000 fois supérieure à celle des mâles, et le sex-ratio est observable dès l’éclosion des œufs.
Capsules d'œufs pondues par une femelle calmar
Embryologie
Les œufs de céphalopodes présentent une grande variété de tailles, de 1 à 30 mm de diamètre. L' ovule fécondé se divise initialement pour former un disque de cellules germinales à un pôle, tandis que le vitellus demeure au pôle opposé. Le disque germinatif se développe pour envelopper et finalement absorber le vitellus, formant ainsi l'embryon. Les tentacules et les bras apparaissent d'abord à l'extrémité postérieure du corps, là où se situerait le pied chez les autres mollusques, et ne migrent vers la tête que plus tard.
L'entonnoir des céphalopodes se développe sur le dessus de leur tête, tandis que la bouche se développe sur la surface opposée. Les premiers stades embryonnaires rappellent ceux des gastéropodes ancestraux et des Monoplacophora actuels .
La coquille se développe à partir de l'ectoderme sous forme d'une structure organique qui se minéralise ensuite. Chez Sepia , qui possède une coquille interne, l'ectoderme forme une invagination dont le pore est scellé avant le dépôt de cette structure organique.
Développement
La durée d'incubation est très variable ; les œufs plus petits, pondus dans des eaux plus chaudes, éclosent plus rapidement, et les nouveau-nés peuvent émerger après seulement quelques jours. Les œufs plus gros, pondus dans des eaux plus froides, peuvent se développer pendant plus d'un an avant d'éclore. Le processus allant de la ponte à l'éclosion suit une trajectoire similaire chez toutes les espèces, la principale variable étant la quantité de vitellus disponible pour les jeunes et le moment de son absorption par l'embryon.
Contrairement à la plupart des autres mollusques, les céphalopodes ne présentent pas de stade larvaire morphologiquement distinct . Leurs juvéniles sont appelés paralarves . Les paralarves ont été observées uniquement chez les Octopodes et les Teuthida (qui constituent la définition moderne des Coleoidea). En revanche, les nautiles nouveau-nés ne sont pas désignés par un terme technique spécifique, car ils ressemblent à des versions miniatures des adultes. Les céphalopodes néonates apprennent rapidement à chasser, perfectionnant leurs stratégies au fil de leurs rencontres avec leurs proies.
La croissance chez les juvéniles est généralement allométrique , tandis que la croissance chez les adultes est isométrique .
Évolution
monoplacophore doté d'une coquille incurvée et effilée , étroitement apparenté aux gastéropodes (escargots) . La ressemblance du céphalopode primitif à coquille, Plectronoceras, avec certains gastéropodes a été invoquée à l'appui de cette théorie. Le développement d'un siphon aurait permis aux coquilles de ces formes primitives de se remplir de gaz (et donc de flotter), assurant ainsi leur maintien et leur verticalité pendant que l'animal rampait sur le sol. Ce siphon aurait également permis de distinguer les véritables céphalopodes d'ancêtres putatifs tels que Knightoconus , dépourvus de siphon . La flottabilité neutre ou positive (c'est-à-dire la capacité de flotter) serait apparue plus tard, suivie par la nage chez les Plectronocerida et, enfin, la propulsion par jet chez les céphalopodes plus évolués
Des restes datant potentiellement du Cambrien inférieur ont été découverts dans la péninsule d'Avalon , correspondant aux données génétiques d'une origine précambrienne. Cependant, il a été démontré par la suite que ce spécimen était un fossile chimérique .
En 2010, certains chercheurs ont proposé que Nectocaris pteryx soit le plus ancien céphalopode connu, dépourvu de coquille et semblant posséder un système de propulsion par jet similaire à celui des céphalopodes « dérivés », ce qui a complexifié la question de l’ordre d’apparition des caractéristiques des céphalopodes. Cependant, la plupart des autres chercheurs doutent que Nectocaris ait été un céphalopode, voire un mollusque, et une étude de 2025 a définitivement établi son lien de parenté avec les chétognathes modernes (vers flèches).
Les premiers céphalopodes étaient probablement des prédateurs situés au sommet de la chaîne alimentaire. Après l' extinction du Cambrien supérieur, qui a entraîné la disparition de nombreux radiodontes , les niches écologiques de prédateurs sont devenues disponibles pour d'autres animaux. Au cours de l'Ordovicien, les céphalopodes primitifs ont connu des phases de diversification pour devenir diversifiés et dominants dans les mers du Paléozoïque et du Mésozoïque .
Au Paléozoïque inférieur, leur répartition était bien plus restreinte ; ils étaient principalement confinés aux zones infralittorales des plateaux continentaux peu profonds des basses latitudes et vivaient généralement en association avec des thrombolites . Un mode de vie plus pélagique s’est progressivement développé au cours de l’Ordovicien. Des céphalopodes d’eaux profondes, bien que rares, ont été découverts dans l’Ordovicien inférieur, mais uniquement dans les eaux de hautes latitudes. L’Ordovicien moyen a vu apparaître les premiers céphalopodes dotés de cloisons suffisamment robustes pour résister aux pressions associées aux eaux profondes et capables de vivre à des profondeurs supérieures à 100-200 m. Le sens d’enroulement de la coquille s’est avéré crucial pour le succès futur des lignées ; l’enroulement endogastrique ne permettait d’atteindre une grande taille qu’avec une coquille droite, tandis que l’enroulement exogastrique – initialement assez rare – a permis le développement des spirales que l’on retrouve dans les archives fossiles, avec leur grande taille et leur diversité respectives. (Endogastrique signifie que la coquille est courbée de sorte que le côté ventral ou inférieur soit concave longitudinalement (abdomen vers l'intérieur) ; exogastrique signifie que la coquille est courbée de sorte que le côté ventral soit convexe longitudinalement (abdomen vers l'extérieur), permettant à l'entonnoir d'être pointé vers l'arrière sous la coquille.)
Un ammonite dont la chambre corporelle est manquante, montrant la surface septale (surtout à droite) avec ses lobes ondulés et ses selles.
Les ancêtres des coléoïdes (dont la plupart des céphalopodes modernes) et ceux du nautile moderne ont divergé au Floien, au début de l'Ordovicien, il y a plus de 470 millions d'années. Les Bactritida , un groupe d'orthocônes du Dévonien-Trias, sont généralement considérés comme paraphylétiques par rapport aux coléoïdes et aux ammonoïdes ; autrement dit, ces derniers groupes seraient issus des Bactritida. Une augmentation de la diversité des coléoïdes et des ammonoïdes est observée au début du Dévonien et correspond à une forte augmentation de la diversité des poissons. Ceci pourrait représenter l'origine des deux groupes dérivés.
Contrairement à la plupart des céphalopodes modernes, la plupart des espèces anciennes possédaient des coquilles protectrices. Ces coquilles étaient d'abord coniques, mais ont ensuite évolué vers des formes nautiloïdes incurvées, comme celles que l'on observe chez les espèces de nautiles modernes .
On pense que la pression compétitive exercée par les poissons a forcé les formes à coquille à migrer vers des eaux plus profondes, ce qui a engendré une pression évolutive favorisant la perte de la coquille et donnant naissance aux coléoïdes modernes. Ce changement a entraîné des coûts métaboliques plus importants liés à la perte de flottabilité, mais leur a permis de recoloniser les eaux peu profondes. Cependant, certains nautiloïdes à coquille droite ont évolué en bélemnites . La perte de la coquille pourrait également résulter d'une pression évolutive visant à accroître la manœuvrabilité, aboutissant à un comportement plus proche de celui des poissons.
L' origine embryologique des appendices des céphalopodes a fait l'objet de débats . Jusqu'au milieu du XXe siècle, l'hypothèse des « bras comme tête » était largement acceptée. Selon cette théorie, les bras et les tentacules des céphalopodes ressemblent aux appendices céphaliques des gastéropodes, suggérant qu'il pourrait s'agir de structures homologues . Les appendices des céphalopodes entourant la bouche, ils pourraient logiquement dériver de tissus céphaliques embryonnaires. Cependant, l'hypothèse des « bras comme pied », proposée par Adolf Naef en 1928, est de plus en plus privilégiée ; par exemple, la cartographie du devenir des bourgeons de membres chez le nautile indique que ces bourgeons proviennent de tissus embryonnaires du « pied ».
Génétique
Le séquençage complet du génome d'un céphalopode demeure un défi pour les chercheurs en raison de la longueur et de la répétition de leur ADN. On a d'abord supposé que les caractéristiques des génomes des céphalopodes résultaient de duplications génomiques complètes . Après le séquençage complet du génome d'une pieuvre à deux points de Californie , celui-ci a présenté des caractéristiques similaires à celles d'autres invertébrés marins, avec des ajouts significatifs au génome, supposés spécifiques aux céphalopodes. Aucune preuve de duplication génomique complète n'a été trouvée.
Le génome de la pieuvre à deux taches de Californie présente des réplications importantes de deux familles de gènes. Il est à noter que ces familles de gènes étendues n'étaient jusqu'alors connues pour présenter un comportement réplicatif que chez les vertébrés. La première famille de gènes identifiée est celle des protocadhérines , impliquées dans le développement neuronal. Les protocadhérines fonctionnent comme des molécules d'adhérence cellulaire, essentielles à la spécificité synaptique . Le mécanisme de réplication de la famille de gènes des protocadhérines chez les vertébrés est attribué à un épissage complexe, ou découpage-collage, à partir d'un locus. Suite au séquençage du génome de la pieuvre à deux taches de Californie, les chercheurs ont découvert que la famille de gènes des protocadhérines chez les céphalopodes s'est étendue dans le génome en raison d'une duplication génique en tandem . Les différents mécanismes de réplication des gènes des protocadhérines indiquent une évolution indépendante de l'expansion de ces gènes chez les vertébrés et les invertébrés. L'analyse des gènes individuels des protocadhérines chez les céphalopodes indique une évolution indépendante entre les espèces de céphalopodes. Une espèce de calmar côtier, *Doryteuthis pealeii*, possédant des familles de gènes de protocadhérine étendues, diffère significativement de celles de la pieuvre à deux taches de Californie, suggérant que l'expansion génique n'a pas eu lieu avant la spéciation chez les céphalopodes. Malgré des mécanismes d'expansion génique différents, les gènes de protocadhérine de la pieuvre à deux taches étaient plus similaires à ceux des vertébrés qu'à ceux du calmar, suggérant un mécanisme d'évolution convergente . La seconde famille de gènes, connue sous le nom de C₂H₂ , code pour de petites protéines fonctionnant comme facteurs de transcription à zinc. Les C₂H₂ sont connus pour moduler les fonctions de l'ADN, de l'ARN et des protéines au sein de la cellule. éléments transposables ainsi qu'une expression de transposons. Bien que le rôle des éléments transposables chez les vertébrés marins demeure relativement méconnu, une expression significative de transposons a été observée dans les tissus du système nerveux . Dans une étude menée sur des vertébrés, l'expression de transposons au cours du développement de la drosophile (Drosophila melanogaster) a activé la diversité génomique entre les neurones . Cette diversité a été associée à une amélioration de la mémoire et de l'apprentissage chez les mammifères. Le lien entre les transposons et l'augmentation des capacités neuronales pourrait éclairer l'intelligence, la mémoire et les fonctions observées chez les céphalopodes
Grâce au séquençage à longues lectures, des chercheurs ont décodé les génomes des céphalopodes et découvert qu'ils étaient profondément remaniés. La comparaison des gènes avec ceux de milliers d'autres espèces a révélé que des groupes de trois gènes ou plus coexistaient chez les calmars et les pieuvres, contrairement à ce qui a été observé chez d'autres animaux. Nombre de ces regroupements se situaient dans le tissu nerveux, suggérant un lien avec l'évolution de leur intelligence.
Phylogénie
Le consensus approximatif de la phylogénie des céphalopodes actuels, d'après Whalen & Landman (2022), est présenté dans le cladogramme . Les taxons minéralisés sont en gras .
La phylogénie interne des céphalopodes est difficile à établir ; de nombreuses techniques moléculaires ont été utilisées, mais les résultats obtenus sont contradictoires. Le nautile est généralement considéré comme un groupe externe, Vampyroteuthis formant un groupe externe par rapport aux autres calmars ; cependant, dans une analyse, les nautiloïdes, les pieuvres et les teuthidés forment une polytomie . Certaines phylogénies moléculaires ne retrouvent pas les coléoïdes minéralisés ( Spirula , Sepia et Metasepia ) comme un clade ; cependant, d’autres retrouvent ce clade, apparemment plus parcimonieux, avec Spirula comme groupe frère de Sepia et Metasepia dans un clade qui avait probablement divergé avant la fin du Trias.
Les estimations moléculaires de la divergence des clades varient. Une estimation « statistiquement robuste » situe la divergence du Nautilus par rapport à celle de la Pieuvre il y a Nautilus chambré ( Nautilus pompilius )Seiche commune ( Sepia officinalis )Bobtail de l'Atlantique ( Sepiola atlantica )Calmar européen ( Loligo vulgaris )Poulpe commun ( Octopus vulgaris )
La classification présentée ici pour les céphalopodes actuels s'inspire largement de la Classification actuelle des céphalopodes actuels (mai 2001), tandis que celle des céphalopodes fossiles reprend les travaux d'Arkell et al. (1957), Teichert et Moore (1964), Teichert (1988) et d'autres auteurs. Les trois sous-classes sont classiques et correspondent aux trois ordres de céphalopodes reconnus par Bather.
Classe des Céphalopodes († indique les groupes éteints )
Sous-classe Nautiloidea : Céphalopodes ectocochliés fondamentaux qui ont fourni la source des Ammonoidea et des Coleoidea.
D'autres classifications diffèrent, principalement quant à la manière dont les différents ordres de décapodes sont apparentés, et quant à savoir s'il s'agit d'ordres ou de familles.
Classification suprafamiliale du Traité de paléontologie des invertébrés
Il s'agit de l'ancienne classification qui combine celles que l'on trouve dans les parties K et L du Traité de paléontologie des invertébrés , qui constitue la base et est en grande partie conservée par les classifications ultérieures.
Les nautiloïdes en général (Teichert et Moore, 1964) séquence comme indiqué.
Sous-classe † Endoceratoidea . Non utilisé par Flower, par exemple Flower et Kummel 1950, les interjocerides inclus dans les Endocerida.
Les révisions ultérieures comprennent l'établissement de trois ordres du Cambrien supérieur, les Plectronocerida, les Protactinocerida et les Yanhecerida ; la séparation des pseudorthocerides en Pseudorthocerida et l'élévation des orthoceratidés en tant que sous-classe des Orthoceratoidea.
Classification de Shevyrev
Shevyrev (2005) a suggéré une division en huit sous-classes, comprenant principalement les formes fossiles les plus diverses et les plus nombreuses, bien que cette classification ait été critiquée comme étant arbitraire, manquant de preuves et basée sur des interprétations erronées d'autres articles.
Fossile pyritisé de Vampyronassa rhodanica , un vampyromorphide du Callovien inférieur ( il y a clades . L'un comprend le nautile et la plupart des nautiloïdes fossiles. L'autre clade ( Neocephalopoda ou Angusteradulata) est plus proche des coléoïdes modernes et inclut les bélemnites, les ammonites et de nombreuses familles d'orthocérides . Il existe également des céphalopodes basaux des Ellesmerocerida traditionnels qui n'appartiennent à aucun des deux clades.
Les coléoïdes, malgré quelques doutes, semblent être monophylétiques d'après les données moléculaires.
Les peuples navigateurs de l'Antiquité connaissaient les céphalopodes, comme en témoigne une sculpture sur pierre découverte lors des fouilles archéologiques minoennes de Knossos (1900-1100 av. J.-C.), en Crète, datant de l'âge du bronze. Cette sculpture représente un pêcheur portant une pieuvre. La Gorgone, créature terrifiante et puissante de la mythologie grecque, pourrait avoir été inspirée par la pieuvre ou le calmar : le corps de la pieuvre représenterait la tête tranchée de Méduse , son bec la langue et les crocs, et ses tentacules les serpents.
Le kraken est un monstre marin légendaire aux proportions gigantesques, censé vivre au large des côtes de Norvège et du Groenland. Il est généralement représenté dans l'art comme un céphalopode géant attaquant des navires. Linné l'a inclus dans la première édition de son Systema Naturae de 1735. Dans un mythe de la création hawaïen , selon lequel le cosmos actuel est le dernier d'une série apparue par étapes à partir des ruines de l'univers précédent, la pieuvre est la seule survivante de cet univers extraterrestre. L' Akkorokamui est un monstre tentaculaire gigantesque issu du folklore aïnou .
Ses nombreux bras qui émanent d'un centre commun signifient que la pieuvre est parfois utilisée pour symboliser une organisation puissante et manipulatrice.