Un transcrit primaire est le produit d'acide ribonucléique ( ARN ) simple brin synthétisé par transcription de l'ADN , puis transformé en divers ARN matures tels que les ARNm , les ARNt et les ARNr . Les transcrits primaires destinés à être des ARNm subissent des modifications en vue de leur traduction . Par exemple, un ARNm précurseur (pré-ARNm) est un type de transcrit primaire qui devient un ARN messager (ARNm) après maturation . L'ARN pré-messager (pré-ARNm) est synthétisé à partir d'une matrice d'ADN dans le noyau cellulaire par transcription . Il constitue la majeure partie de l'ARN nucléaire hétérogène (ARNhn). Une fois sa maturation terminée, le pré-ARNm est appelé « ARN messager mature », ou simplement « ARN messager ». Le terme ARNhn est souvent utilisé comme synonyme de pré-ARNm, bien que, au sens strict, il puisse inclure des transcrits d'ARN nucléaires qui ne deviennent pas des ARNm cytoplasmiques. Plusieurs étapes contribuent à la production des transcrits primaires. Toutes ces étapes impliquent une série d'interactions pour initier et achever la transcription de l'ADN dans le noyau des eucaryotes . Certains facteurs jouent un rôle clé dans l'activation et l'inhibition de la transcription, régulant ainsi la production des transcrits primaires. La transcription produit des transcrits primaires qui sont ensuite modifiés par plusieurs processus, notamment la coiffe 5' , la polyadénylation 3' et l'épissage alternatif . En particulier, l'épissage alternatif contribue directement à la diversité des ARNm présents dans les cellules. Les modifications des transcrits primaires ont fait l'objet d'études approfondies visant à mieux comprendre le rôle et l'importance de ces transcrits. Des études expérimentales basées sur les modifications moléculaires des transcrits primaires et les processus se déroulant avant et après la transcription ont permis une meilleure compréhension des maladies impliquant ces transcrits.![]()
Chez les eucaryotes, trois types d'ARN — l'ARNr , l'ARNt et l'ARNm — sont produits grâce à l'activité de trois ARN polymérases distinctes, tandis que chez les procaryotes , une seule ARN polymérase est nécessaire à la synthèse de tous les types de molécules d'ARN. L'ARN polymérase II des eucaryotes transcrit le transcrit primaire, destiné à être transformé en ARNm, à partir de la matrice d'ADN antisens dans le sens 5' → 3'. Ce transcrit primaire nouvellement synthétisé est complémentaire du brin antisens de l'ADN. L'ARN polymérase II construit le transcrit primaire en utilisant quatre résidus de ribonucléosides monophosphates spécifiques ( adénosine monophosphate (AMP), cytidine monophosphate (CMP), guanosine monophosphate (GMP) et uridine monophosphate (UMP)) qui sont ajoutés successivement au groupe hydroxyle 3' de l'extrémité 3' de l'ARNm en cours de synthèse.
Des études sur les transcrits primaires produits par l'ARN polymérase II révèlent qu'un transcrit primaire moyen mesure 7 000 nucléotides , certains pouvant atteindre 20 000 nucléotides. L'inclusion de séquences exoniques et introniques dans les transcrits primaires explique la différence de taille entre ces transcrits plus longs et les ARNm matures plus courts, prêts à être traduits en protéines.activateurs . Les facteurs de transcription , protéines se liant à des éléments d'ADN pour activer ou réprimer la transcription, se fixent aux activateurs et recrutent des enzymes qui modifient la composition des nucléosomes , rendant ainsi l'ADN plus ou moins accessible à l'ARN polymérase. Les combinaisons spécifiques de facteurs de transcription activateurs ou inhibiteurs se liant aux régions d'ADN activatrices déterminent si le gène avec lequel l'activateur interagit est activé ou non pour la transcription. L'activation de la transcription dépend de la capacité du complexe d'élongation de la transcription, lui-même constitué de divers facteurs de transcription, à induire la dissociation de l'ARN polymérase du complexe Médiateur qui relie une région activatrice au promoteur.
L’inhibition de l’activité de l’ARN polymérase peut également être régulée par des séquences d’ADN appelées silencieux . À l’instar des activateurs, les silencieux peuvent être situés en amont ou en aval des gènes qu’ils régulent. Ces séquences d’ADN se lient à des facteurs qui contribuent à la déstabilisation du complexe d’initiation nécessaire à l’activation de l’ARN polymérase, et inhibent ainsi la transcription.
La modification des histones par les facteurs de transcription est un autre facteur de régulation clé de la transcription par l'ARN polymérase. En général, les facteurs qui induisent l'acétylation des histones activent la transcription, tandis que ceux qui induisent leur désacétylation l'inhibent . L'acétylation des histones induit une répulsion entre les composants négatifs au sein des nucléosomes, permettant ainsi l'accès de l'ARN polymérase. La désacétylation des histones stabilise les nucléosomes compacts, inhibant l'accès de l'ARN polymérase. Outre les profils d'acétylation des histones, les profils de méthylation des régions promotrices de l'ADN peuvent réguler l'accès de l'ARN polymérase à une matrice donnée. L'ARN polymérase est souvent incapable de synthétiser un transcrit primaire si la région promotrice du gène cible contient des cytosines méthylées spécifiques – des résidus qui entravent la liaison des facteurs d'activation de la transcription et recrutent d'autres enzymes pour stabiliser une structure nucléosomale compacte, empêchant ainsi l'accès de l'ARN polymérase et la production de transcrits primaires
Boucles R
Les boucles R se forment lors de la transcription. Une boucle R est une structure d'acide nucléique à trois brins contenant une région hybride ADN-ARN et un brin d'ADN simple brin non matrice associé. Les régions d' ADN activement transcrites forment souvent des boucles R vulnérables aux dommages à l'ADN . Les introns réduisent la formation de boucles R et les dommages à l'ADN dans les gènes de levure fortement exprimés.
Stress de transcription
Des lésions de l'ADN surviennent quotidiennement dans chaque cellule, leur nombre pouvant atteindre plusieurs dizaines, voire centaines de milliers par cellule. Ces lésions peuvent entraver la transcription primaire. L' expression des gènes elle-même est une source de lésions endogènes de l'ADN, du fait de la fragilité de l'ADN simple brin. D'autres sources de lésions de l'ADN incluent les conflits entre le complexe de transcription primaire et le complexe de réplication de l'ADN , ainsi que l'activité de certaines enzymes comme les topoisomérases et les enzymes de réparation par excision de base . Bien que ces processus soient étroitement régulés et généralement précis, des erreurs peuvent occasionnellement se produire, entraînant des cassures de l'ADN qui provoquent des réarrangements chromosomiques ou la mort cellulaire .
Traitement de l'ARN
Polyadénylation
Épissage alternatif
Dans les cellules eucaryotes complexes, un seul transcrit primaire peut générer de grandes quantités d'ARNm matures grâce à l'épissage alternatif. Ce processus est régulé de sorte que chaque ARNm mature puisse coder pour plusieurs protéines.
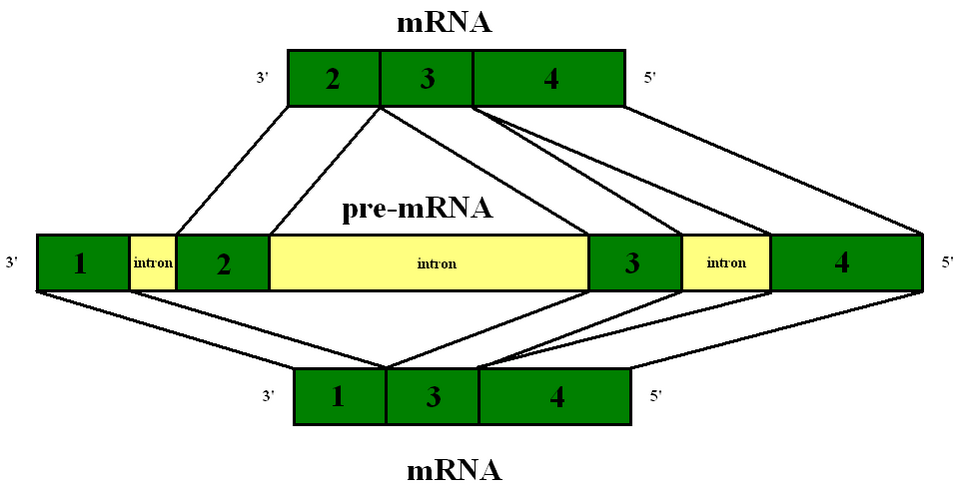
L'effet de l'épissage alternatif sur l'expression des gènes est observable chez les eucaryotes complexes, qui possèdent un nombre fixe de gènes dans leur génome mais produisent un nombre beaucoup plus important de produits géniques différents. La plupart des transcrits d'ARN pré-messager eucaryotes contiennent de multiples introns et exons. Les diverses combinaisons possibles des sites d'épissage 5' et 3' dans un ARN pré-messager peuvent conduire à différentes excisions et combinaisons d'exons, tandis que les introns sont éliminés de l'ARN messager mature. Ainsi, différents types d'ARN messagers matures sont générés. L'épissage alternatif a lieu dans un grand complexe protéique appelé spliceosome . Il est crucial pour la régulation tissulaire et développementale de l'expression des gènes. L'épissage alternatif peut être affecté par divers facteurs, notamment des mutations telles que les translocations chromosomiques .
Chez les procaryotes, l'épissage s'effectue par clivage autocatalytique ou par clivage endolytique. Les clivages autocatalytiques, qui n'impliquent aucune protéine, concernent généralement les séquences codant pour l'ARNr, tandis que les clivages endolytiques correspondent aux précurseurs de l'ARNt.
Recherche
L’exposition des cellules KB résistantes au méthotrexate au 5- fluorouracile (FUra) a entraîné une réduction de moitié des niveaux d’ARNm de la dihydrofolate réductase (DHFR) totale, tandis que le niveau de pré-ARNm de la DHFR contenant certains introns est resté inchangé. La demi-vie de l’ARNm ou du pré-ARNm de la DHFR n’a pas varié significativement, mais le taux de transition de l’ARN de la DHFR du noyau vers le cytoplasme a diminué, suggérant que le FUra pourrait influencer la maturation de l’ARNm et/ou la stabilité nucléaire de l’ARNm de la DHFR.
Chez Drosophila et Aedes , la taille des ARNhn (pré-ARNm) était plus importante chez Aedes en raison de son génome plus grand, bien que les deux espèces produisent des ARNm matures de taille et de complexité de séquence similaires. Ceci indique que la taille des ARNhn augmente avec la taille du génome.
Dans les cellules HeLa , des groupes de spliceosomes se forment sur le pré-ARNm au sein de granules nucléaires ; cette formation est dépendante de la température et influencée par des séquences d’ARN spécifiques. Le ciblage du pré-ARNm et le chargement des facteurs d’épissage dans les granules sont essentiels à la formation des groupes de spliceosomes, ce qui donne un aspect granuleux.
Le recrutement de l'ARN pré-messager dans les granules nucléaires a augmenté de manière significative l'efficacité de l'épissage et les niveaux de protéines, indiquant que la proximité des granules améliore l'efficacité de l'épissage.
Maladies apparentées
La recherche a également permis d'approfondir nos connaissances sur certaines maladies liées à des modifications des transcrits primaires. Une étude a porté sur les récepteurs d'œstrogènes et l'épissage alternatif. L'article intitulé « Épissage alternatif du transcrit primaire du récepteur alpha des œstrogènes humains : mécanismes de saut d'exons », par Paola Ferro, Alessandra Forlani, Marco Muselli et Ulrich Pfeffer du laboratoire d'oncologie moléculaire de l'Institut national de recherche sur le cancer de Gênes, en Italie, explique que 1 785 nucléotides de la région de l'ADN codant pour le récepteur alpha des œstrogènes (ER-alpha) sont répartis sur une région contenant plus de 300 000 nucléotides dans le transcrit primaire. L'épissage de ce pré-ARNm conduit fréquemment à des variants ou à différents types d'ARNm dépourvus d'un ou plusieurs exons ou régions nécessaires au codage des protéines. Ces variants ont été associés à la progression du cancer du sein . Dans le cycle de vie des rétrovirus , l'ADN proviral est incorporé à la transcription de l'ADN de la cellule infectée. Les rétrovirus doivent convertir leur pré-ARNm en ADN pour que ce dernier puisse s'intégrer à l'ADN de la cellule hôte qu'ils infectent. La formation de cette matrice d'ADN est donc une étape cruciale pour leur réplication. Le type cellulaire, sa différenciation ou son état modifié, ainsi que son état physiologique, induisent des variations importantes dans la disponibilité et l'activité de certains facteurs nécessaires à la transcription. Ces variables engendrent une grande diversité d'expression des gènes viraux. Par exemple, les cellules en culture produisant activement des virions infectieux de virus de la leucémie aviaire ou murine (ASLV ou MLV) contiennent des taux d'ARN viral si élevés que 5 à 10 % de l'ARNm d'une cellule peuvent être d'origine virale. Ceci démontre que les transcrits primaires produits par ces rétrovirus ne suivent pas toujours la voie classique de la synthèse protéique et ne se reconvertissent pas en ADN pour se multiplier et se propager.