Un ARN non codant ( ARNnc ) est une molécule d'ARN fonctionnelle qui n'est pas traduite en protéine . La séquence d'ADN à partir de laquelle un ARN non codant fonctionnel est transcrit est souvent appelée gène d'ARN . Les types d'ARN non codants abondants et fonctionnellement importants comprennent les ARN de transfert (ARNt) et les ARN ribosomiques (ARNr), ainsi que les petits ARN tels que les microARN , les siARN , les piARN , les snoARN , les snARN , les exARN , les scaARN et les longs ARNnc tels que Xist et HOTAIR .
Le nombre d'ARN non codants dans le génome humain est inconnu ; cependant, des études transcriptomiques et bioinformatiques récentes suggèrent qu'il existe des milliers de transcriptions non codantes. De nombreux ARN non codants nouvellement identifiés ont des fonctions inconnues, voire inexistantes. Il n'y a pas de consensus sur la part de transcription non codante qui est fonctionnelle : certains pensent que la plupart des ARN non codants sont des « ARN indésirables » non fonctionnels, des transcriptions fausses, tandis que d'autres s'attendent à ce que de nombreuses transcriptions non codantes aient des fonctions à découvrir.
Histoire et découverte
Les acides nucléiques ont été découverts pour la première fois en 1868 par Friedrich Miescher , et en 1939, l'ARN était impliqué dans la synthèse des protéines . Deux décennies plus tard, Francis Crick a prédit un composant d'ARN fonctionnel qui médiatise la traduction ; il a estimé que l'ARN est mieux adapté à l'appariement de bases avec un transcrit d'ARNm qu'un polypeptide pur .
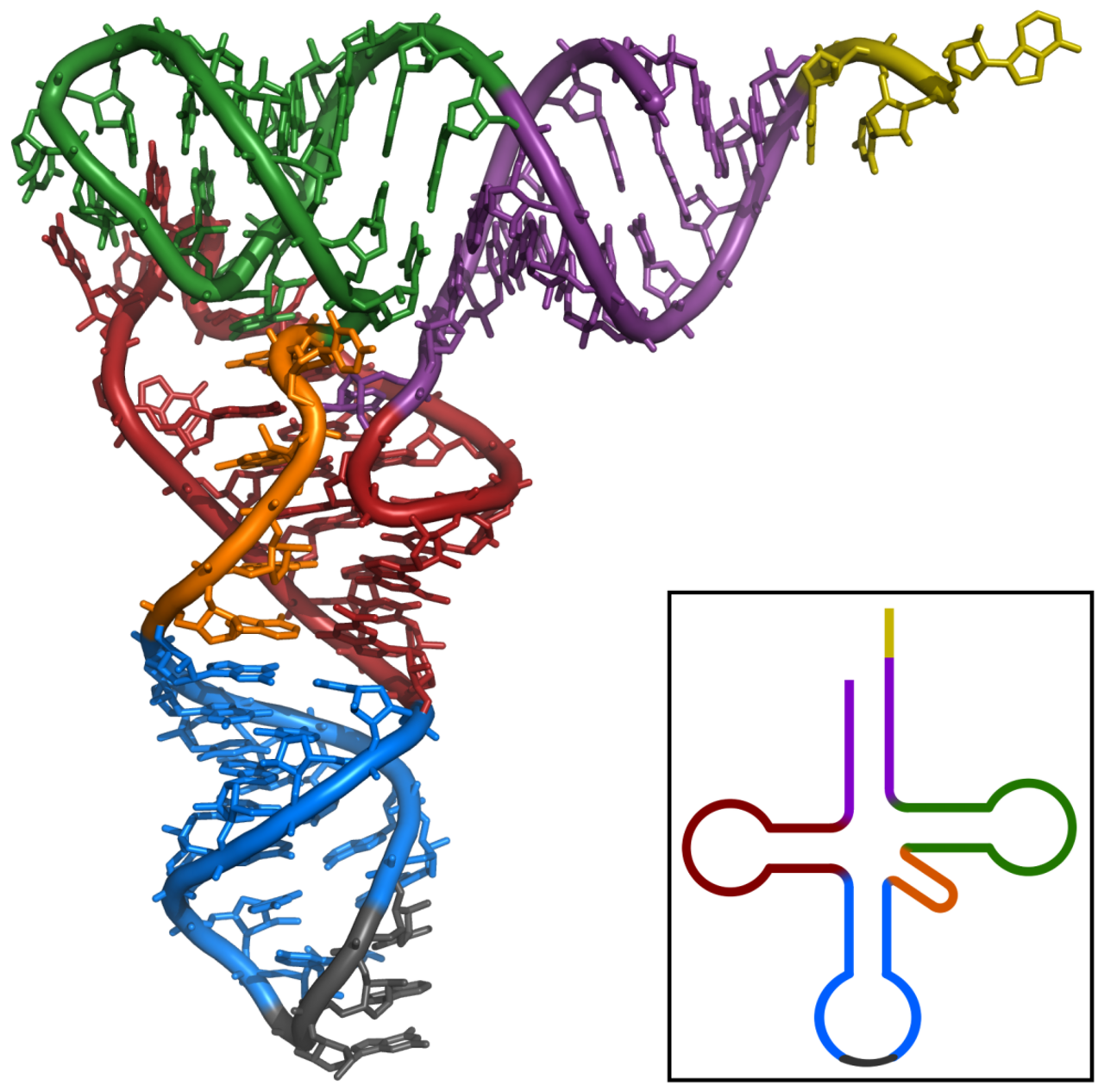
Français Le premier ARN non codant à être caractérisé était un ARNt d'alanine trouvé dans la levure de boulanger , sa structure a été publiée en 1965. Pour produire un échantillon d'ARNt d'alanine purifié, Robert W. Holley et al. ont utilisé 140 kg de levure de boulanger commerciale pour donner seulement 1 g d'ARNt Ala purifié pour analyse. L'ARNt de 80 nucléotides a été séquencé en étant d'abord digéré par la ribonucléase pancréatique (produisant des fragments se terminant par Cytosine ou Uridine ) puis par la ribonucléase takadiastase Tl (produisant des fragments se terminant par Guanosine ). La chromatographie et l'identification des extrémités 5' et 3' ont ensuite aidé à organiser les fragments pour établir la séquence d'ARN. Des trois structures initialement proposées pour cet ARNt, la structure en « feuille de trèfle » a été proposée indépendamment dans plusieurs publications ultérieures. La structure secondaire en trèfle a été finalisée après une analyse par cristallographie aux rayons X réalisée par deux groupes de recherche indépendants en 1974.
L'ARN ribosomique a été le suivant à être découvert, suivi par l'ARNr au début des années 1980. Depuis lors, la découverte de nouveaux ARN non codants s'est poursuivie avec les snoRNA , Xist , CRISPR et bien d'autres. Les ajouts notables récents incluent les riboswitches et les miRNA ; la découverte du mécanisme d'ARNi associé à ce dernier a valu à Craig C. Mello et Andrew Fire le prix Nobel de physiologie ou médecine 2006.
Les découvertes récentes d’ARNnc ont été réalisées grâce à des méthodes expérimentales et bioinformatiques .
Rôles biologiques
Les ARN non codants appartiennent à plusieurs groupes et sont impliqués dans de nombreux processus cellulaires. Ceux-ci vont des ARN non codants d'importance centrale qui sont conservés dans toute ou la plupart des cellules vivantes jusqu'aux ARN non codants plus transitoires spécifiques à une ou quelques espèces étroitement apparentées. On pense que les ARN non codants les plus conservés sont des fossiles moléculaires ou des reliques du dernier ancêtre commun universel et du monde de l'ARN , et leurs rôles actuels restent principalement dans la régulation du flux d'informations de l'ADN vers les protéines.
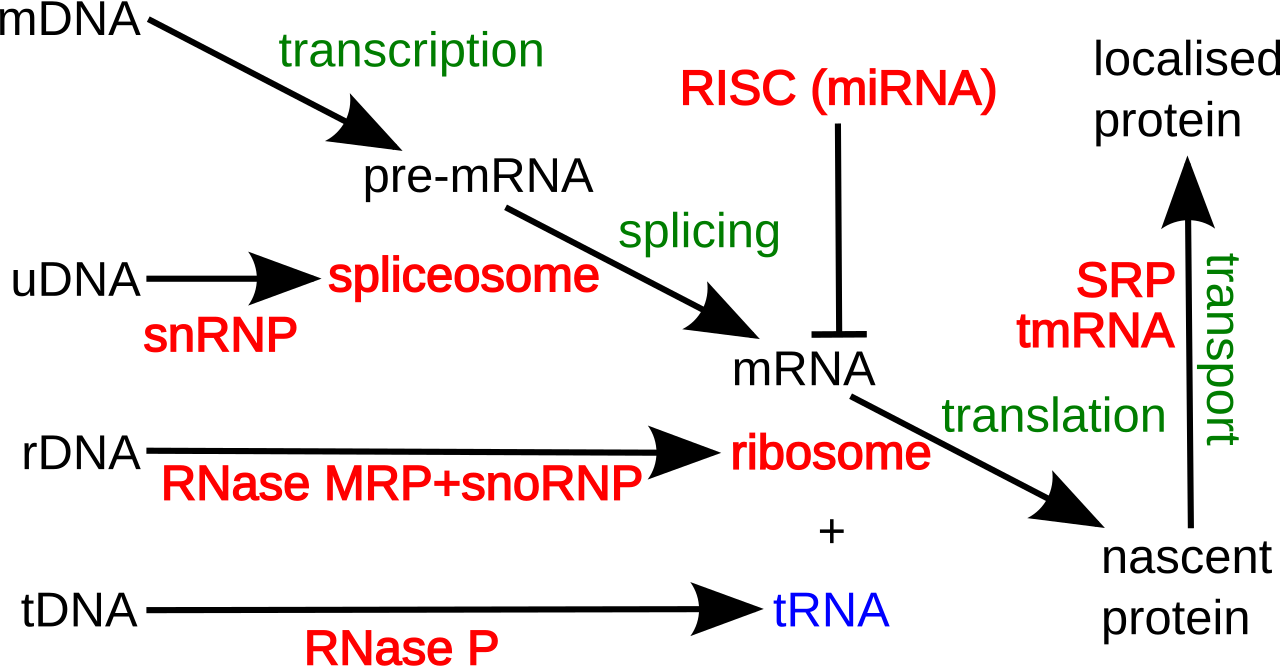
En traduction
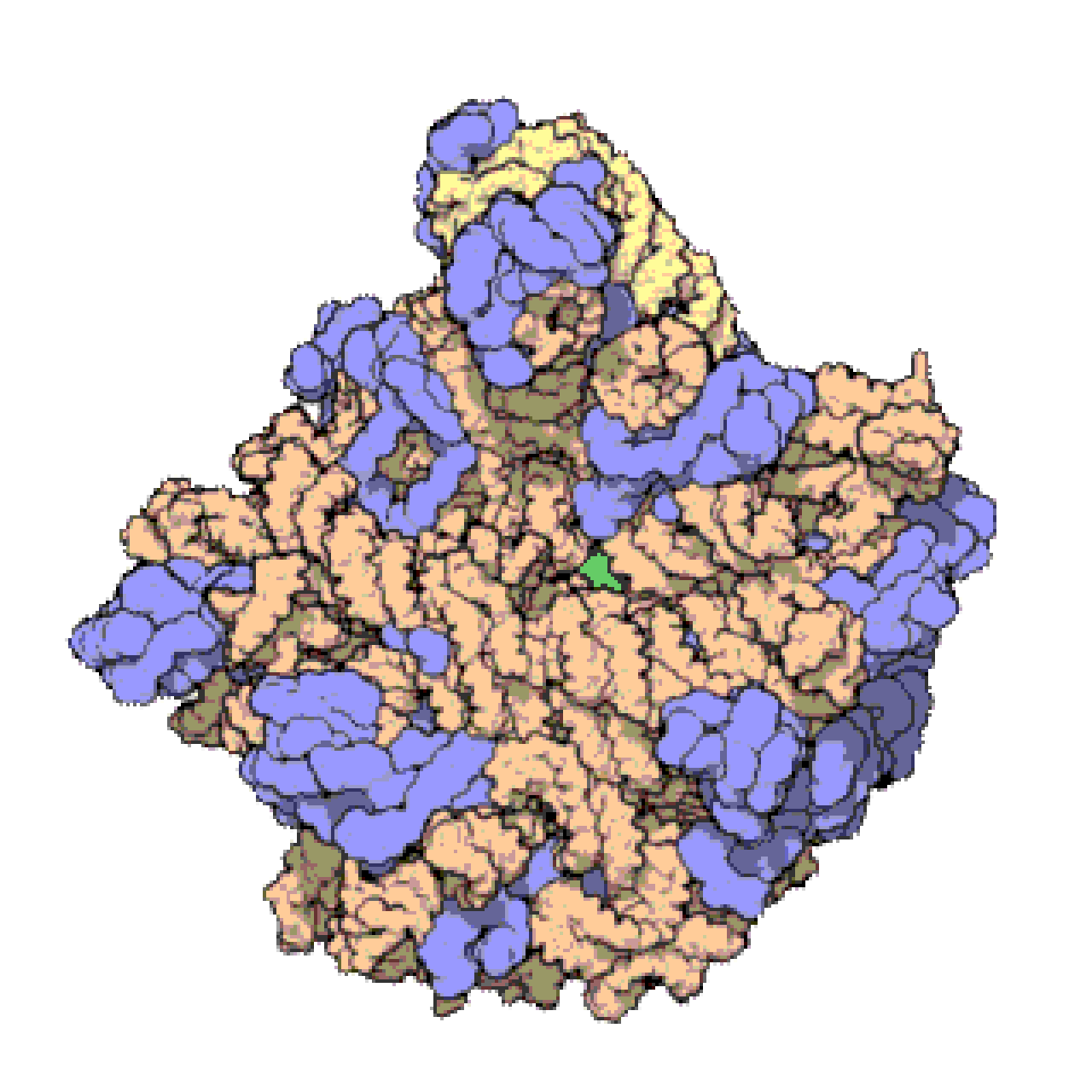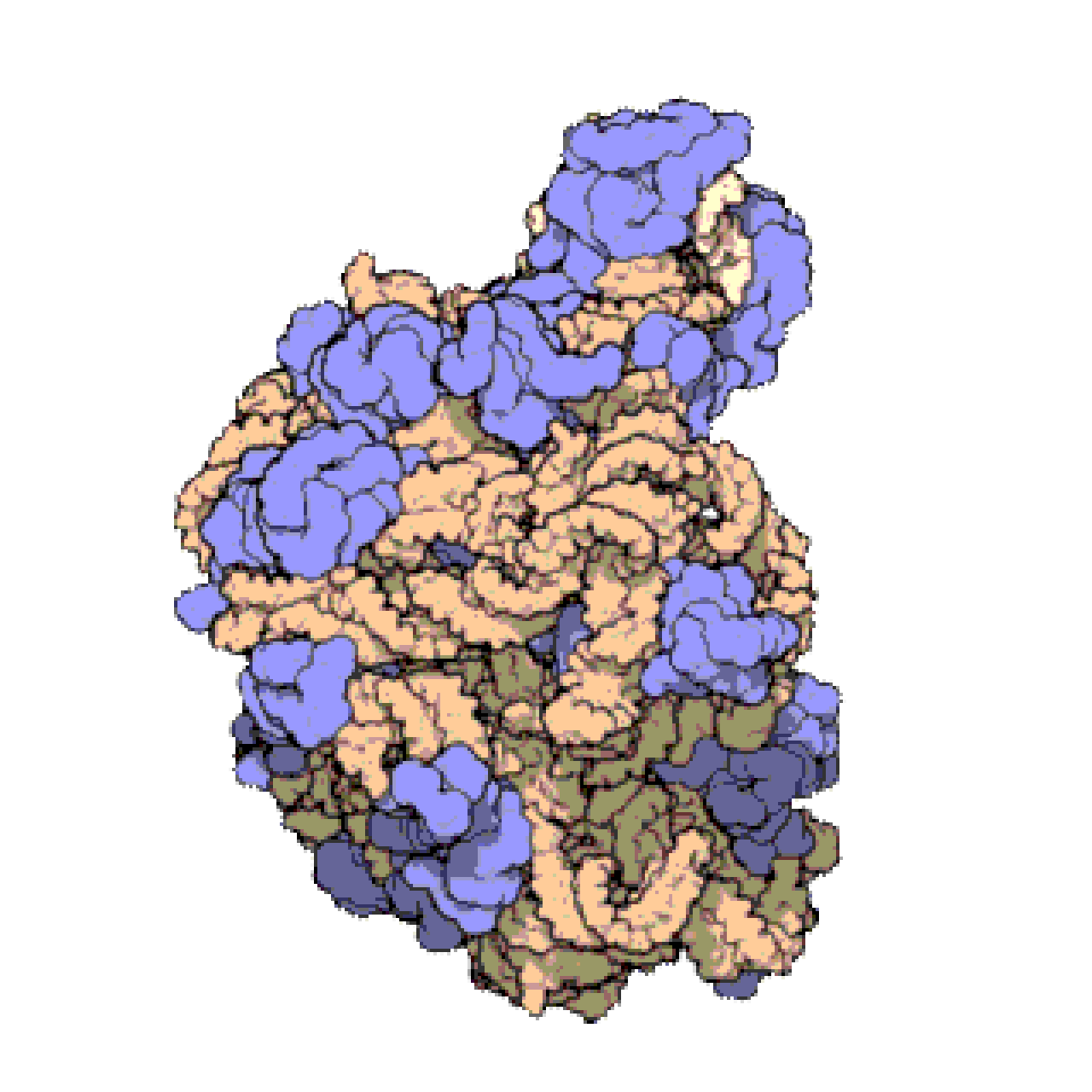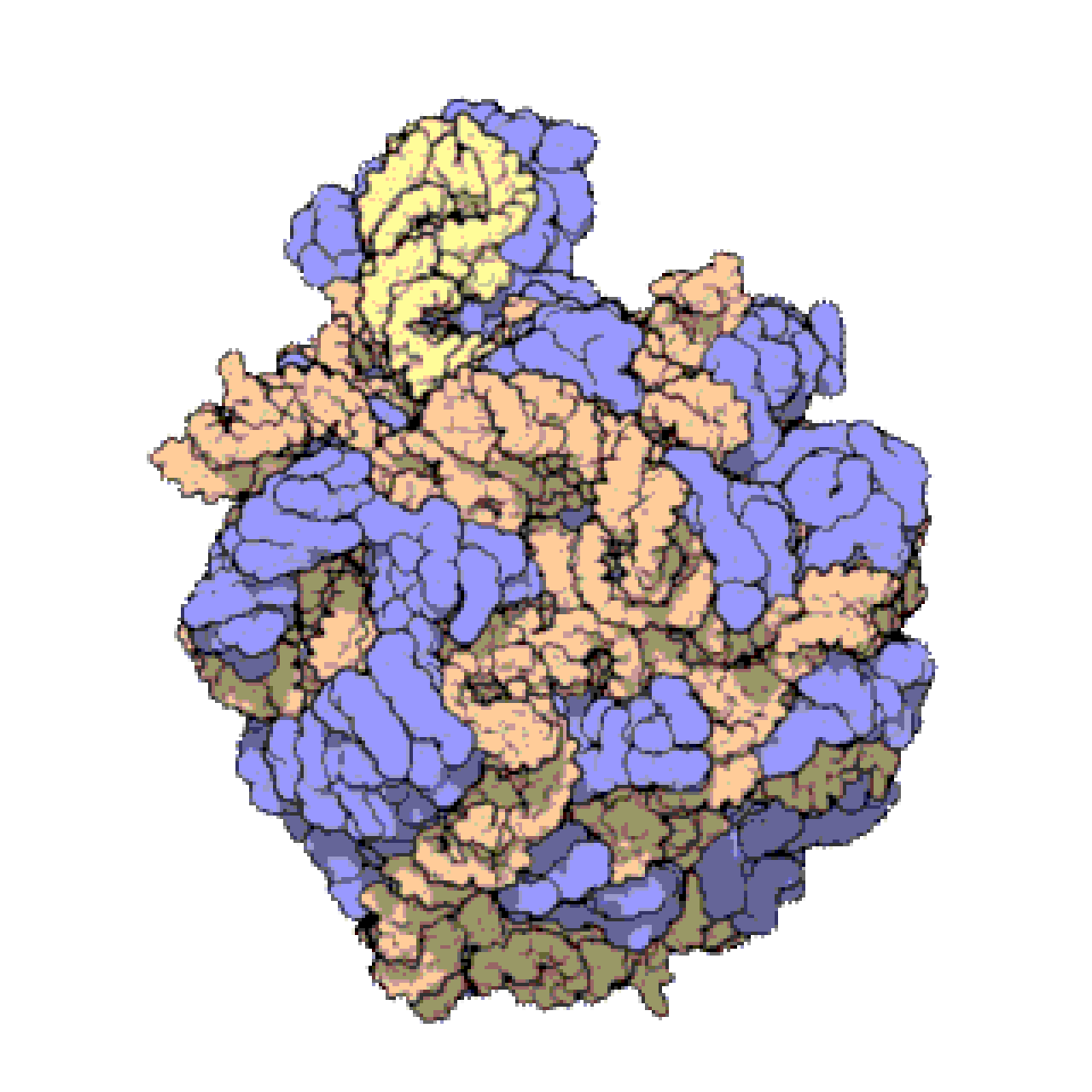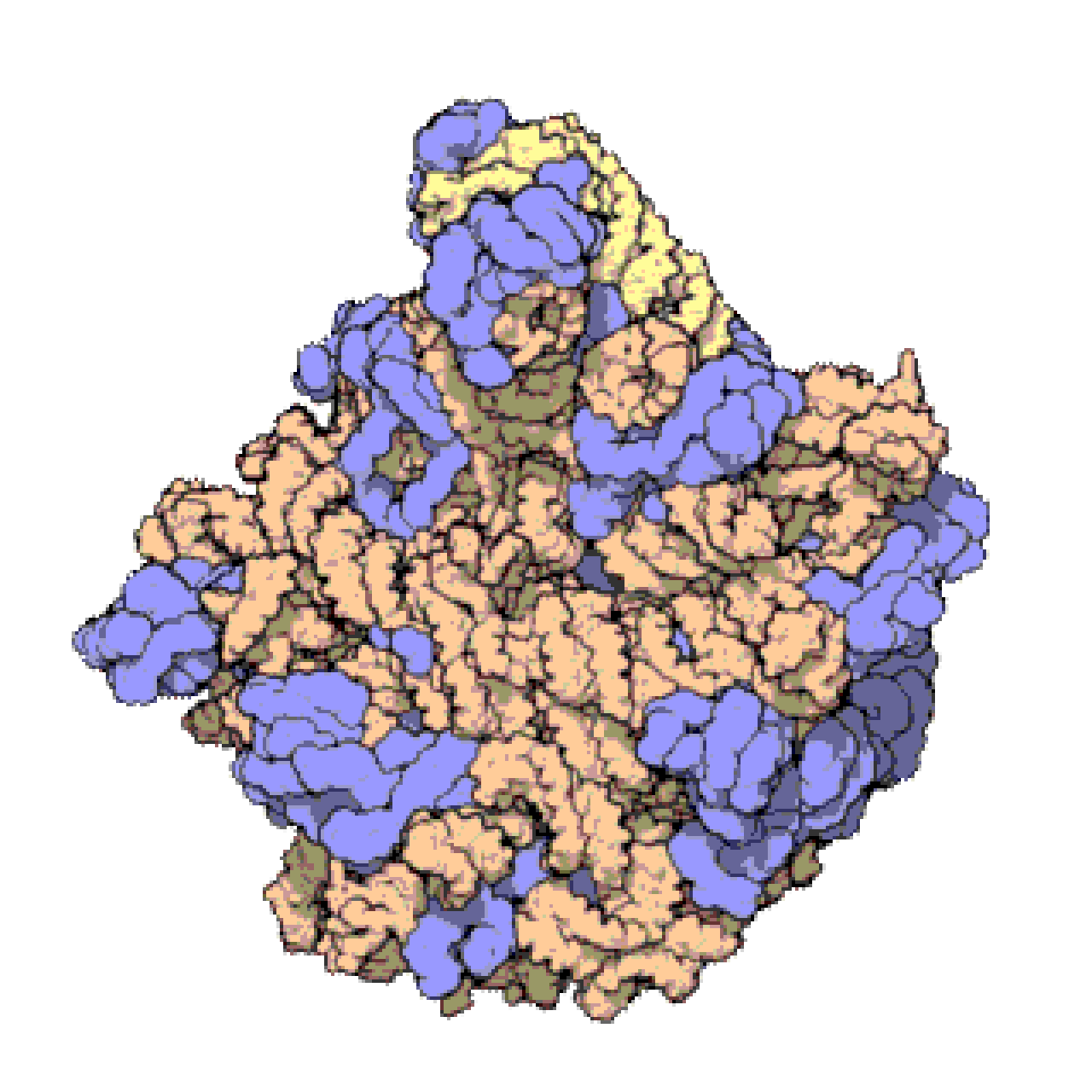
Français De nombreux ARNnc conservés, essentiels et abondants sont impliqués dans la traduction . Les particules de ribonucléoprotéines (RNP) appelées ribosomes sont les « usines » où la traduction a lieu dans la cellule. Le ribosome est constitué de plus de 60 % d'ARN ribosomique ; ceux-ci sont constitués de 3 ARNnc chez les procaryotes et de 4 ARNnc chez les eucaryotes . Les ARN ribosomiques catalysent la traduction des séquences nucléotidiques en protéines. Un autre ensemble d'ARNnc, les ARN de transfert , forment une « molécule adaptatrice » entre l'ARNm et la protéine. Les ARNnc H/ACA box et C/D box sont des ARNnc trouvés dans les archées et les eucaryotes. La RNase MRP est limitée aux eucaryotes. Les deux groupes d'ARNnc sont impliqués dans la maturation de l'ARNr. Les ARNnc guident les modifications covalentes de l'ARNr, de l'ARNt et des ARNnc ; La RNase MRP clive l' espaceur interne transcrit 1 entre les ARNr 18S et 5,8S. L'ARNnc omniprésent, la RNase P , est un parent évolutif de la RNase MRP. La RNase P fait mûrir les séquences d'ARNt en générant des extrémités 5' matures des ARNt en clivant les éléments leaders 5' des ARNt précurseurs. Une autre RNP omniprésente appelée SRP reconnaît et transporte des protéines naissantes spécifiques vers le réticulum endoplasmique chez les eucaryotes et la membrane plasmique chez les procaryotes . Chez les bactéries, l'ARN messager de transfert (ARNtm) est un RNP impliqué dans le sauvetage des ribosomes bloqués, le marquage des polypeptides incomplets et la promotion de la dégradation de l'ARNm aberrant.
Dans l'épissage de l'ARN
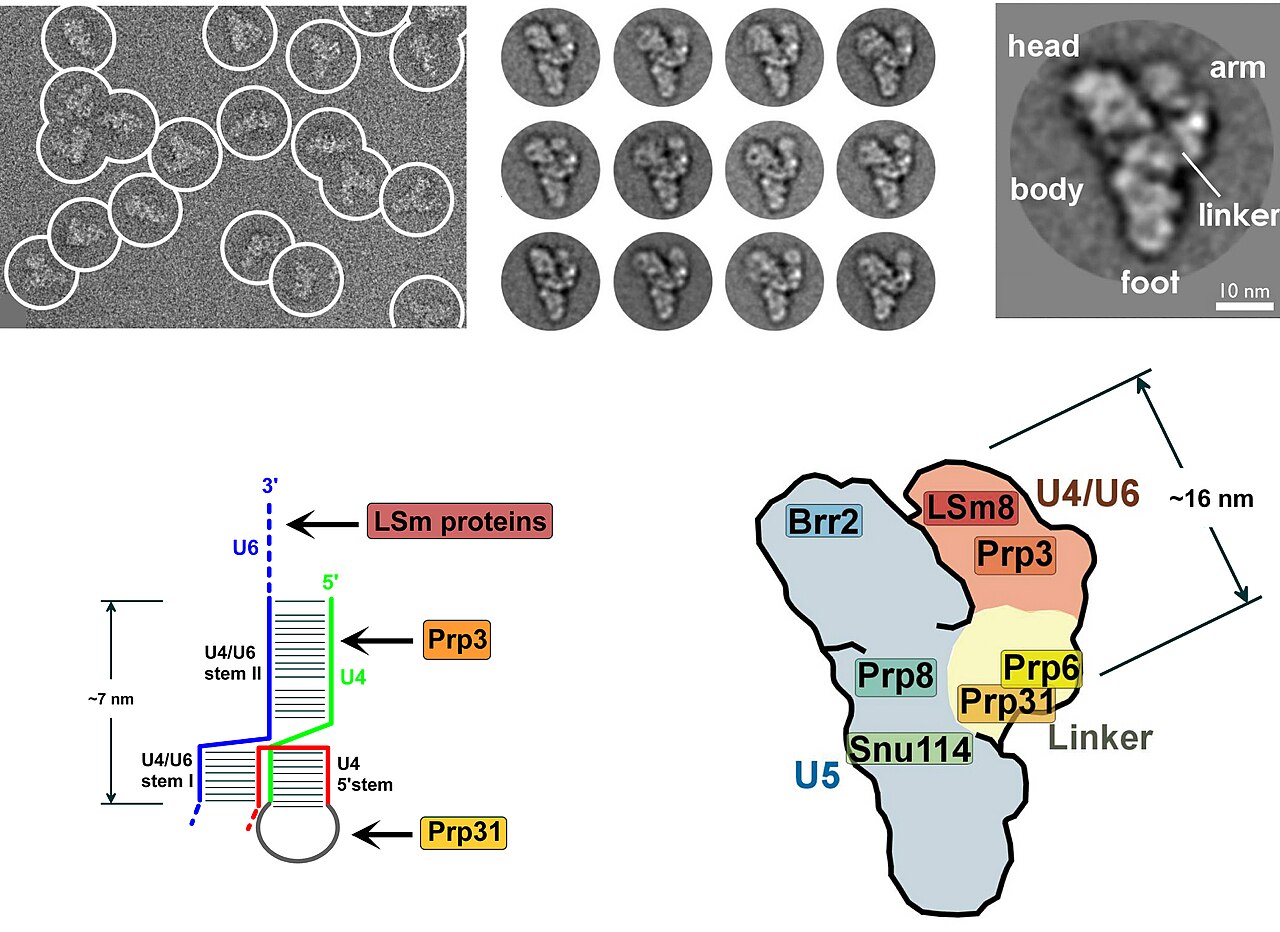
Chez les eucaryotes, le spliceosome effectue les réactions d'épissage essentielles pour éliminer les séquences d'introns , ce processus est nécessaire à la formation d' ARNm mature . Le spliceosome est un autre RNP souvent connu sous le nom de snRNP ou tri-snRNP. Il existe deux formes différentes du spliceosome, les formes majeure et mineure. Les composants ncRNA du spliceosome majeur sont U1 , U2 , U4 , U5 et U6 . Les composants ncRNA du spliceosome mineur sont U11 , U12 , U5 , U4atac et U6atac .
Un autre groupe d'introns peut catalyser leur propre élimination des transcrits de l'hôte ; on les appelle ARN auto-épissés. Il existe deux groupes principaux d'ARN auto-épissés : les introns catalytiques du groupe I et les introns catalytiques du groupe II . Ces ARNnc catalysent leur propre excision des précurseurs d'ARNm, d'ARNt et d'ARNr dans un large éventail d'organismes.
Chez les mammifères, il a été découvert que les snoRNA peuvent également réguler l' épissage alternatif de l'ARNm, par exemple le snoRNA HBII-52 régule l'épissage du récepteur de la sérotonine 2C .
Chez les nématodes, l' ARNnc SmY semble être impliqué dans l'épissage trans de l'ARNm .
Dans la réplication de l'ADN
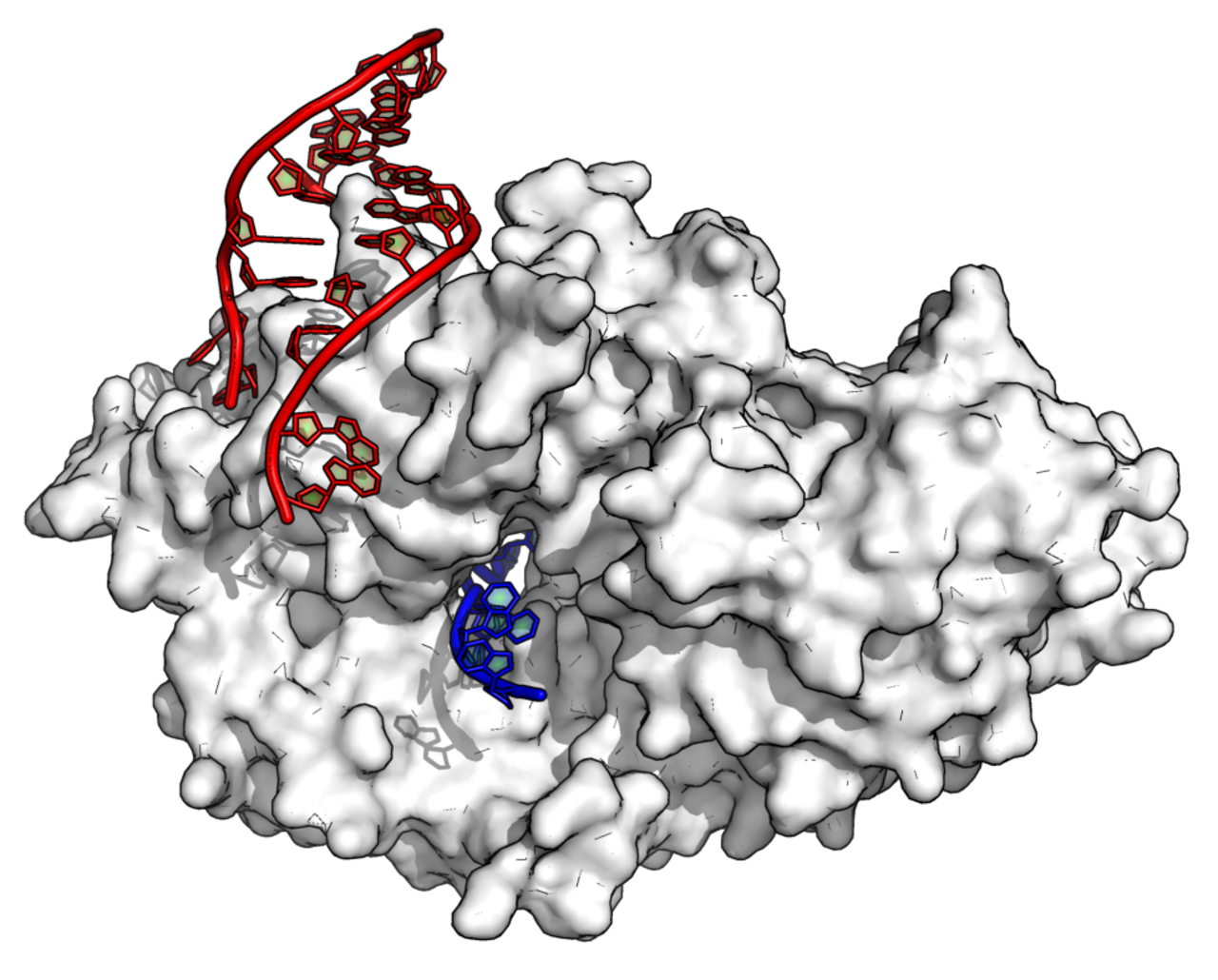
Les ARN Y sont des tiges-boucles, nécessaires à la réplication de l'ADN par le biais d'interactions avec la chromatine et les protéines d'initiation (y compris le complexe de reconnaissance d'origine ). Ils sont également des composants de la particule ribonucléoprotéique Ro60 qui est une cible des anticorps auto-immuns chez les patients atteints de lupus érythémateux disséminé .
Dans la régulation des gènes
L' expression de plusieurs milliers de gènes est régulée par les ARNnc. Cette régulation peut se produire en trans ou en cis . Il existe de plus en plus de preuves qu'un type particulier d'ARNnc appelé ARN enhancer , transcrit à partir de la région enhancer d'un gène, agit pour favoriser l'expression des gènes.
Transaction
Chez les eucaryotes supérieurs, les microARN régulent l'expression des gènes. Un seul miARN peut réduire les niveaux d'expression de centaines de gènes. Le mécanisme par lequel les molécules de miARN matures agissent est la complémentarité partielle avec une ou plusieurs molécules d'ARN messager (ARNm), généralement dans les 3' UTR . La principale fonction des miARN est de réguler à la baisse l'expression des gènes.
Il a également été démontré que la RNase P de l'ARNnc influence l'expression des gènes. Dans le noyau humain, la RNase P est nécessaire à la transcription normale et efficace de divers ARNnc transcrits par l'ARN polymérase III . Il s'agit notamment des gènes d'ARNt, d'ARNr 5S , d'ARN SRP et d'ARNsn U6 . La RNase P exerce son rôle dans la transcription par association avec Pol III et la chromatine des gènes actifs d'ARNt et d'ARNr 5S.
Il a été démontré que l'ARN 7SK , un ARNnc métazoaire , agit comme un régulateur négatif du facteur d'élongation de l' ARN polymérase II P-TEFb , et que cette activité est influencée par les voies de réponse au stress.
L'ARNnc bactérien, l'ARN 6S , s'associe spécifiquement à l'holoenzyme ARN polymérase contenant le facteur de spécificité sigma70 . Cette interaction réprime l'expression d'un promoteur dépendant de sigma70 pendant la phase stationnaire .
Un autre ARNnc bactérien, l'ARN OxyS, réprime la traduction en se liant aux séquences Shine-Dalgarno, bloquant ainsi la liaison aux ribosomes. L'ARN OxyS est induit en réponse au stress oxydatif chez Escherichia coli.
L'ARN B2 est un petit transcrit non codant de l'ARN polymérase III qui réprime la transcription de l'ARNm en réponse au choc thermique dans les cellules de souris. L'ARN B2 inhibe la transcription en se liant au noyau Pol II. Grâce à cette interaction, l'ARN B2 s'assemble en complexes de préinitiation au niveau du promoteur et bloque la synthèse de l'ARN.
Une étude récente a montré que le simple fait de transcrire une séquence d'ARNnc peut avoir une influence sur l'expression des gènes. La transcription des ARNnc par l'ARN polymérase II est nécessaire au remodelage de la chromatine chez Schizosaccharomyces pombe . La chromatine est progressivement convertie en une configuration ouverte, à mesure que plusieurs espèces d'ARNnc sont transcrites.
Cis-agissant
Un certain nombre d'ARNnc sont intégrés dans les régions non traduites ( UTR ) 5' des gènes codant pour des protéines et influencent leur expression de diverses manières. Par exemple, un riboswitch peut se lier directement à une petite molécule cible ; la liaison de la cible affecte l'activité du gène.
Les séquences de leader d'ARN se trouvent en amont du premier gène des opérons biosynthétiques d'acides aminés. Ces éléments d'ARN forment l'une des deux structures possibles dans les régions codant des séquences peptidiques très courtes qui sont riches en acide aminé produit final de l'opéron. Une structure de terminateur se forme lorsqu'il y a un excès de l'acide aminé régulateur et que le mouvement du ribosome sur le transcrit leader n'est pas entravé. Lorsqu'il y a une déficience de l'ARNt chargé de l'acide aminé régulateur, le ribosome traduisant le peptide leader se bloque et la structure antiterminatrice se forme. Cela permet à l'ARN polymérase de transcrire l'opéron. Les leaders d'ARN connus sont le leader de l'opéron histidine , le leader de l'opéron leucine , le leader de l'opéron thréonine et le leader de l'opéron tryptophane .
Les éléments de réponse au fer (IRE) sont liés aux protéines de réponse au fer (IRP). L'IRE se trouve dans les UTR de divers ARNm dont les produits sont impliqués dans le métabolisme du fer . Lorsque la concentration en fer est faible, les IRP se lient à l'ARNm de la ferritine IRE, ce qui entraîne une répression de la traduction.
Les sites d'entrée internes des ribosomes (IRES) sont des structures d'ARN qui permettent l' initiation de la traduction au milieu d'une séquence d'ARNm dans le cadre du processus de synthèse des protéines .
En défense du génome
Les ARN interagissant avec Piwi (piRNA) exprimés dans les testicules et les cellules somatiques des mammifères forment des complexes ARN-protéine avec les protéines Piwi . Ces complexes piRNA (piRC) ont été liés au silençage génique transcriptionnel des rétrotransposons et d'autres éléments génétiques dans les cellules germinales , en particulier celles de la spermatogenèse .
Les répétitions palindromiques courtes groupées et régulièrement espacées (CRISPR) sont des répétitions présentes dans l' ADN de nombreuses bactéries et archées . Les répétitions sont séparées par des espaceurs de longueur similaire. Il a été démontré que ces espaceurs peuvent être dérivés de phages et contribuer par la suite à protéger la cellule contre l'infection.
Structure des chromosomes
La télomérase est une enzyme RNP qui ajoute des répétitions de séquences d'ADN spécifiques (« TTAGGG » chez les vertébrés) aux régions télomériques , qui se trouvent aux extrémités des chromosomes eucaryotes . Les télomères contiennent du matériel d'ADN condensé, ce qui confère une stabilité aux chromosomes. L'enzyme est une transcriptase inverse qui transporte l'ARN télomérase , qui est utilisé comme modèle lorsqu'il allonge les télomères, qui sont raccourcis après chaque cycle de réplication .
Xist (X-inactive-specific transcript) est un long gène ncRNA sur le chromosome X des mammifères placentaires qui agit comme effecteur majeur du processus d'inactivation du chromosome X formant les corps de Barr . Un ARN antisens , Tsix , est un régulateur négatif de Xist. Les chromosomes X dépourvus d'expression de Tsix (et ayant donc des niveaux élevés de transcription Xist) sont inactivés plus fréquemment que les chromosomes normaux. Chez les drosophiles , qui utilisent également un système de détermination du sexe XY , les ARN roX (ARN sur le X) sont impliqués dans la compensation de dosage. Xist et roX fonctionnent tous deux par régulation épigénétique de la transcription via le recrutement d' enzymes modifiant les histones .
ARN bifonctionnel
Les ARN bifonctionnels , ou ARN à double fonction , sont des ARN qui ont deux fonctions distinctes. La majorité des ARN bifonctionnels connus sont des ARNm qui codent à la fois une protéine et des ARNnc. Cependant, un nombre croissant d'ARNnc appartiennent à deux catégories différentes d'ARNnc ; par exemple, les ARNsno à boîte H/ACA et les ARNmi .
Deux exemples bien connus d'ARN bifonctionnels sont l'ARN SgrS et l'ARNIII . Cependant, on connaît l'existence d'une poignée d'autres ARN bifonctionnels (par exemple, l'activateur du récepteur des stéroïdes/SRA, l'ARN VegT, [48 l'ARN Oskar, ENOD40 , l'ARN p53 l'ARN SR1 , et l'ARN Spot 42. ) . Les ARN bifonctionnels ont fait l'objet d'un numéro spécial de Biochimie en 2011.
En tant qu'hormone
Il existe un lien important entre certains ARN non codants et le contrôle des voies régulées par les hormones. Chez la drosophile , des hormones telles que l'ecdysone et l'hormone juvénile peuvent favoriser l'expression de certains miARN. De plus, cette régulation se produit à des moments temporels distincts au cours du développement de Caenorhabditis elegans . Chez les mammifères, le miR-206 est un régulateur crucial du récepteur alpha des œstrogènes .
Les ARN non codants sont essentiels au développement de plusieurs organes endocriniens, ainsi qu'aux maladies endocriniennes telles que le diabète sucré . Plus précisément, dans la lignée cellulaire MCF-7, l'ajout de 17β- estradiol a augmenté la transcription globale des ARN non codants appelés lncRNA à proximité des gènes codants activés par les œstrogènes.
En évitement pathogène
Il a été démontré que C. elegans apprend et hérite de l'évitement pathogène après exposition à un seul ARN non codant d'un pathogène bactérien .
Rôles dans la maladie
Comme pour les protéines , les mutations ou les déséquilibres dans le répertoire d’ARNnc dans le corps peuvent provoquer diverses maladies.
Cancer
De nombreux ARNnc présentent des profils d'expression anormaux dans les tissus cancéreux . Il s'agit notamment des miARN , des ARNnc longs de type ARNm , GAS5 , SNORD50 , ARN télomérase et ARN Y. [ Les miARN sont impliqués dans la régulation à grande échelle de nombreux gènes codant pour des protéines, les ARN Y sont importants pour l'initiation de la réplication de l'ADN, ARN télomérase qui sert d'amorce à la télomérase, une RNP qui étend les régions télomériques aux extrémités des chromosomes (voir télomères et maladie pour plus d'informations). La fonction directe des ARNnc longs de type ARNm est moins claire.
Il a été démontré que les mutations germinales dans les précurseurs primaires miR-16-1 et miR-15 sont beaucoup plus fréquentes chez les patients atteints de leucémie lymphoïde chronique que dans les populations témoins.
Il a été suggéré qu'un SNP rare (rs11614913) qui chevauche hsa-mir-196a-2 a été trouvé associé au carcinome pulmonaire non à petites cellules . De même, un criblage de 17 miRNA qui ont été prédits pour réguler un certain nombre de gènes associés au cancer du sein a trouvé des variations dans les microRNA miR-17 et miR-30c-1 des patients ; ces patients n'étaient pas porteurs de mutations BRCA1 ou BRCA2 , ce qui laisse penser que le cancer du sein familial peut être causé par une variation de ces miRNA. Le suppresseur de tumeur p53 est sans doute l'agent le plus important dans la prévention de la formation et de la progression des tumeurs. La protéine p53 fonctionne comme un facteur de transcription avec un rôle crucial dans l'orchestration de la réponse au stress cellulaire. En plus de son rôle crucial dans le cancer, p53 a été impliqué dans d'autres maladies, notamment le diabète, la mort cellulaire après ischémie et diverses maladies neurodégénératives telles que Huntington, Parkinson et Alzheimer. Des études ont suggéré que l’expression de p53 est soumise à la régulation par l’ARN non codant.
Français Un autre exemple d'ARN non codant dérégulé dans les cellules cancéreuses est le long ARN non codant Linc00707. Linc00707 est régulé à la hausse et éponge les miARN dans les cellules souches mésenchymateuses dérivées de la moelle osseuse humaine, le cancer gastrique ou le cancer du sein, et favorise ainsi l'ostéogenèse, contribue à la progression du carcinome hépatocellulaire, favorise la prolifération et les métastases, ou régule indirectement l'expression des protéines impliquées dans l'agressivité du cancer, respectivement.
Syndrome de Prader-Willi
Il a été démontré que la suppression des 48 copies de la boîte C/D du snoRNA SNORD116 est la principale cause du syndrome de Prader-Willi . Le syndrome de Prader-Willi est un trouble du développement associé à une suralimentation et à des difficultés d'apprentissage. Le SNORD116 a des sites cibles potentiels dans un certain nombre de gènes codant pour des protéines et pourrait jouer un rôle dans la régulation de l'épissage alternatif.
Autisme
Le locus chromosomique contenant le groupe de gènes SNORD115 du petit ARN nucléolaire a été dupliqué chez environ 5 % des individus présentant des traits autistiques . Un modèle de souris conçu pour avoir une duplication du groupe SNORD115 présente un comportement de type autistique. Une petite étude récente sur des tissus cérébraux post-mortem a démontré une expression altérée des longs ARN non codants dans le cortex préfrontal et le cervelet des cerveaux autistes par rapport aux témoins.
Hypoplasie cartilage-cheveux
Il a été démontré que des mutations au sein de la RNase MRP provoquent une hypoplasie cartilage-cheveux , une maladie associée à un ensemble de symptômes tels qu'une petite taille, des cheveux clairsemés, des anomalies squelettiques et un système immunitaire affaibli, fréquente chez les Amish et les Finlandais . transition A vers G au niveau du nucléotide 70 qui se trouve dans une région de boucle à deux bases 5' d'un pseudo-nœud conservé . Cependant, de nombreuses autres mutations au sein de la RNase MRP provoquent également une CHH.
La maladie d'Alzheimer
L'ARN antisens BACE1-AS est transcrit à partir du brin opposé à BACE1 et est régulé à la hausse chez les patients atteints de la maladie d'Alzheimer . BACE1-AS régule l'expression de BACE1 en augmentant la stabilité de l'ARNm de BACE1 et en générant davantage de BACE1 par un mécanisme de rétroaction post-transcriptionnelle. Par le même mécanisme, il augmente également les concentrations de bêta-amyloïde , le principal constituant des plaques séniles. Les concentrations de BACE1-AS sont élevées chez les sujets atteints de la maladie d'Alzheimer et chez les souris transgéniques porteuses de la protéine précurseur de l'amyloïde.
miR-96 et perte auditive
La variation dans la région de la graine du miR-96 mature a été associée à une perte auditive progressive autosomique dominante chez les humains et les souris. Les souris mutantes homozygotes étaient profondément sourdes, ne montrant aucune réponse cochléaire . Les souris hétérozygotes et les humains perdent progressivement la capacité d'entendre.
ARN de transfert mitochondrial
Un certain nombre de mutations dans les ARNt mitochondriaux ont été liées à des maladies telles que le syndrome MELAS , le syndrome MERRF et l'ophtalmoplégie externe progressive chronique .
Distinction entre ARN fonctionnel (ARNf) et ARNnc
Les scientifiques ont commencé à distinguer l'ARN fonctionnel ( ARNf ) de l'ARNnc, pour décrire les régions fonctionnelles au niveau de l'ARN qui peuvent ou non être des transcrits d'ARN autonomes. Cela implique que l'ARNf (tel que les riboswitches, les éléments SECIS et d'autres régions cis-régulatrices) n'est pas l'ARNnc. Pourtant, l'ARNf pourrait également inclure l'ARNm , car il s'agit d'ARN codant pour une protéine, et est donc fonctionnel. De plus, les ARN évolués artificiellement relèvent également du terme générique d'ARNf. Certaines publications indiquent que l'ARNnc et l'ARNf sont presque synonymes, mais d'autres ont souligné qu'une grande proportion d'ARNnc annotés n'ont probablement aucune fonction. Il a également été suggéré d'utiliser simplement le terme ARN , puisque la distinction avec un ARN codant pour une protéine ( ARN messager ) est déjà donnée par le qualificatif ARNm . Cela élimine l'ambiguïté lorsqu'on s'adresse à un gène « codant pour un ARN non codant ». En outre, il peut y avoir un certain nombre d'ARNnc qui sont mal notés dans la littérature publiée et dans les ensembles de données.