Les neurones sont les principaux constituants du tissu nerveux chez tous les animaux , à l'exception des éponges et des placozoaires . Les plantes et les champignons sont dépourvus de cellules nerveuses. Des données moléculaires suggèrent que la capacité à générer des signaux électriques est apparue il y a environ 700 à 800 millions d'années, durant le Tonien . Les précurseurs des neurones étaient les cellules sécrétrices peptidergiques . Celles-ci ont acquis progressivement de nouveaux modules génétiques leur permettant de créer des structures postsynaptiques et des canaux ioniques générant des signaux électriques rapides. Cette capacité à générer des signaux électriques a constitué une innovation majeure dans l'évolution du système nerveux.
Les neurones sont généralement classés en trois types selon leur fonction. Les neurones sensoriels réagissent à des stimuli tels que le toucher, le son ou la lumière qui affectent les cellules des organes sensoriels ; ils envoient des signaux à la moelle épinière , puis à l’aire sensorielle du cerveau . Les neurones moteurs reçoivent des signaux du cerveau et de la moelle épinière pour contrôler diverses fonctions, allant des contractions musculaires à l’activité glandulaire . Les interneurones connectent les neurones entre eux au sein d’une même région du cerveau ou de la moelle épinière. Lorsque plusieurs neurones sont fonctionnellement connectés, ils forment un circuit neuronal .
Un neurone possède toutes les structures des autres cellules, telles qu'un noyau , des mitochondries et un appareil de Golgi , mais aussi des structures uniques comme l' axone et les dendrites . Le soma , ou corps cellulaire, est une structure compacte, et l'axone et les dendrites sont des filaments qui en émergent. Les dendrites se ramifient généralement abondamment et s'étendent sur plusieurs centaines de micromètres à partir du soma. L'axone quitte le soma au niveau d'un renflement appelé cône d'émergence et parcourt jusqu'à un mètre chez l'humain, voire plus chez d'autres espèces. Il se ramifie, mais conserve généralement un diamètre constant. À l'extrémité des ramifications de l'axone se trouvent les terminaisons axonales , où le neurone peut transmettre un signal à une autre cellule à travers la synapse . Certains neurones sont dépourvus de dendrites ou d'axone. Le terme « neurite » désigne indifféremment une dendrite ou un axone, notamment lorsque la cellule est indifférenciée .
La plupart des neurones reçoivent des signaux via leurs dendrites et leur soma, puis les transmettent le long de leur axone. Au niveau de la majorité des synapses, les signaux passent de l'axone d'un neurone à la dendrite d'un autre. Cependant, les synapses peuvent connecter deux axones ou deux dendrites. La transmission des signaux est à la fois électrique et chimique. Les neurones sont électriquement excitables grâce au maintien de gradients de potentiel membranaire . Si le potentiel varie suffisamment sur un court intervalle, le neurone génère une impulsion électrochimique de type « tout ou rien » appelée potentiel d'action . Ce potentiel se propage rapidement le long de l'axone et active les connexions synaptiques qu'il atteint. Les signaux synaptiques peuvent être excitateurs ou inhibiteurs , augmentant ou diminuant le potentiel de membrane au niveau du soma.
Dans la plupart des cas, les neurones sont générés par des cellules souches neurales pendant le développement cérébral et l'enfance. La neurogenèse cesse en grande partie à l'âge adulte dans la plupart des régions du cerveau.
système nerveux , avec les cellules gliales qui leur assurent un soutien structurel et métabolique. Le système nerveux est composé du système nerveux central , qui comprend le cerveau et la moelle épinière , et du système nerveux périphérique , qui comprend les systèmes nerveux autonome , entérique et somatique . Chez les vertébrés, la majorité des neurones appartiennent au système nerveux central, mais certains se trouvent dans les ganglions périphériques , et de nombreux neurones sensoriels sont situés dans des organes sensoriels tels que la rétine et la cochlée .Dans le système nerveux périphérique, les axones peuvent s'assembler en faisceaux nerveux qui constituent les nerfs (à l'image des fils d'un câble). Dans le système nerveux central, ces faisceaux d'axones sont appelés voies nerveuses .
Anatomie et histologie


Les neurones sont hautement spécialisés dans le traitement et la transmission des signaux cellulaires. Compte tenu de la diversité des fonctions assurées dans les différentes parties du système nerveux, leur forme, leur taille et leurs propriétés électrochimiques présentent une grande variété. Par exemple, le diamètre du soma d'un neurone peut varier de 4 à 100 micromètres .
- Le soma est le corps du neurone. Comme il contient le noyau , la plupart des synthèses protéiques s'y déroulent. Le noyau peut avoir un diamètre de 3 à 18 micromètres.
- Les dendrites d'un neurone sont des prolongements cellulaires très ramifiés. Cette forme et cette structure sont métaphoriquement comparées à un arbre dendritique. Les ramifications forment des motifs fractals qui se répètent à différentes échelles. C'est au sein de cet arbre fractal que la majorité des informations entrant dans le neurone transitent par l' épine dendritique .
- L' axone est une projection fine, semblable à un câble, dont la longueur peut atteindre des dizaines, des centaines, voire des dizaines de milliers de fois le diamètre du soma. Il transporte principalement les signaux nerveux depuis le soma et certains types d'informations vers celui-ci. De nombreux neurones ne possèdent qu'un seul axone, mais celui-ci peut – et le fait généralement – se ramifier abondamment, permettant ainsi la communication avec de nombreuses cellules cibles. La partie de l'axone où il émerge du soma est appelée cône d'émergence . Outre son rôle anatomique, le cône d'émergence présente également la plus forte densité de canaux sodiques voltage-dépendants . Cela en fait la partie la plus facilement excitable du neurone et la zone d'initiation du potentiel d'action. En termes électrophysiologiques, il possède le potentiel de seuil le plus négatif .
- Bien que l'axone et le cône d'émergence soient généralement impliqués dans la transmission de l'information, cette région peut également recevoir des informations provenant d'autres neurones.
- La terminaison axonale se situe à l'extrémité de l'axone la plus éloignée du soma et contient des synapses . Les boutons synaptiques sont des structures spécialisées où sont libérés des neurotransmetteurs pour communiquer avec les neurones cibles. Outre les boutons synaptiques présents à la terminaison axonale, un neurone peut posséder des boutons en passant , répartis le long de l'axone.
La conception communément admise du neurone attribue des fonctions spécifiques à ses différents composants anatomiques ; cependant, les dendrites et les axones agissent souvent de manière contraire à leur fonction dite principale.motoneurone humain peut dépasser un mètre de long, s'étendant de la base de la colonne vertébrale jusqu'aux orteils.
Les neurones sensoriels peuvent posséder des axones qui s'étendent des orteils jusqu'à la colonne postérieure de la moelle épinière, sur plus de 1,5 mètre chez l'adulte. Les girafes possèdent des axones uniques de plusieurs mètres de long qui parcourent toute la longueur de leur cou. La plupart des connaissances sur la fonction axonale proviennent de l'étude de l' axone géant du calmar , un modèle expérimental idéal en raison de sa taille relativement importante (0,5 à 1 millimètre d'épaisseur, plusieurs centimètres de long).
Les neurones pleinement différenciés sont définitivement post-mitotiques Cependant, les cellules souches présentes dans le cerveau adulte peuvent régénérer des neurones fonctionnels tout au long de la vie de l'organisme (voir neurogenèse ). Les astrocytes sont des cellules gliales en forme d'étoile qui peuvent se différencier en neurones grâce à leur pluripotence , une caractéristique similaire à celle des cellules souches
Membrane
Comme toutes les cellules animales, le corps cellulaire de chaque neurone est entouré d'une membrane plasmique , une bicouche de molécules lipidiques contenant de nombreuses structures protéiques. Une bicouche lipidique est un excellent isolant électrique , mais dans les neurones, plusieurs de ces structures protéiques sont électriquement actives. Parmi celles-ci figurent les canaux ioniques , qui permettent le passage des ions chargés électriquement à travers la membrane, et les pompes ioniques, qui transportent chimiquement les ions d'un côté à l'autre de la membrane. La plupart des canaux ioniques sont de type « porte » , c'est-à-dire perméables uniquement à certains types d'ions. Certains canaux ioniques sont voltage-dépendants : leur ouverture et leur fermeture peuvent être commutées par modification du potentiel de membrane. D'autres sont chimiodépendants : leur ouverture et leur fermeture peuvent être commutées par interaction avec des substances chimiques diffusant dans le liquide extracellulaire. Ces ions comprennent le sodium , le potassium , le chlorure et le calcium . Les interactions entre les canaux ioniques et les pompes ioniques génèrent une différence de potentiel transmembranaire, généralement inférieure à 0,1 volt à l'état basal. Ce potentiel remplit deux fonctions : d'une part, il alimente divers complexes protéiques voltage-dépendants intégrés à la membrane ; d'autre part, il permet la transmission de signaux électriques entre différentes parties de la membrane.
Histologie et structure interne
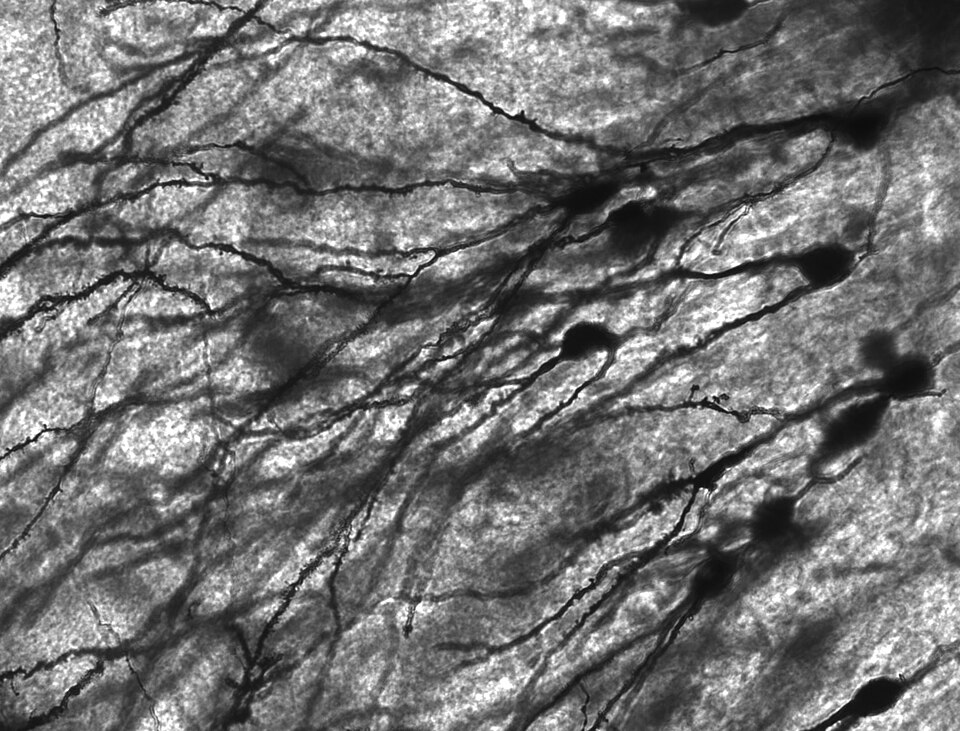

De nombreux amas microscopiques appelés corps de Nissl (ou substance de Nissl) sont observés lorsque les corps cellulaires des neurones sont colorés avec un colorant basophile. Ces structures sont constituées de réticulum endoplasmique rugueux et d'ARN ribosomique associé . Nommées d'après le psychiatre et neuropathologiste allemand Franz Nissl (1860-1919), elles interviennent dans la synthèse protéique et leur présence s'explique par l'intense activité métabolique des neurones. Les colorants basophiles tels que l'aniline ou (faiblement) l'hématoxyline mettent en évidence les composants chargés négativement et se lient ainsi au squelette phosphate de l'ARN ribosomique.
Le corps cellulaire d'un neurone est soutenu par un réseau complexe de protéines structurales appelées neurofilaments , qui, associés aux neurotubules (microtubules neuronaux), s'assemblent en neurofibrilles plus larges. Certains neurones contiennent également des granules pigmentaires, tels que la neuromélanine (un pigment brun-noir, sous-produit de la synthèse des catécholamines ) et la lipofuscine (un pigment brun-jaune), dont l'accumulation augmente avec l'âge. Parmi les autres protéines structurales importantes pour la fonction neuronale figurent l'actine et la tubuline des microtubules . La β-tubuline de classe III est présente presque exclusivement dans les neurones. L'actine se trouve principalement à l'extrémité des axones et des dendrites au cours du développement neuronal. À ce niveau, sa dynamique peut être modulée par une interaction avec les microtubules.
Les axones et les dendrites présentent des caractéristiques structurales internes différentes. Les axones typiques contiennent rarement des ribosomes , sauf dans leur segment initial. Les dendrites contiennent du réticulum endoplasmique granuleux ou des ribosomes, en quantité décroissante à mesure qu'elles s'éloignent du corps cellulaire.
Classification
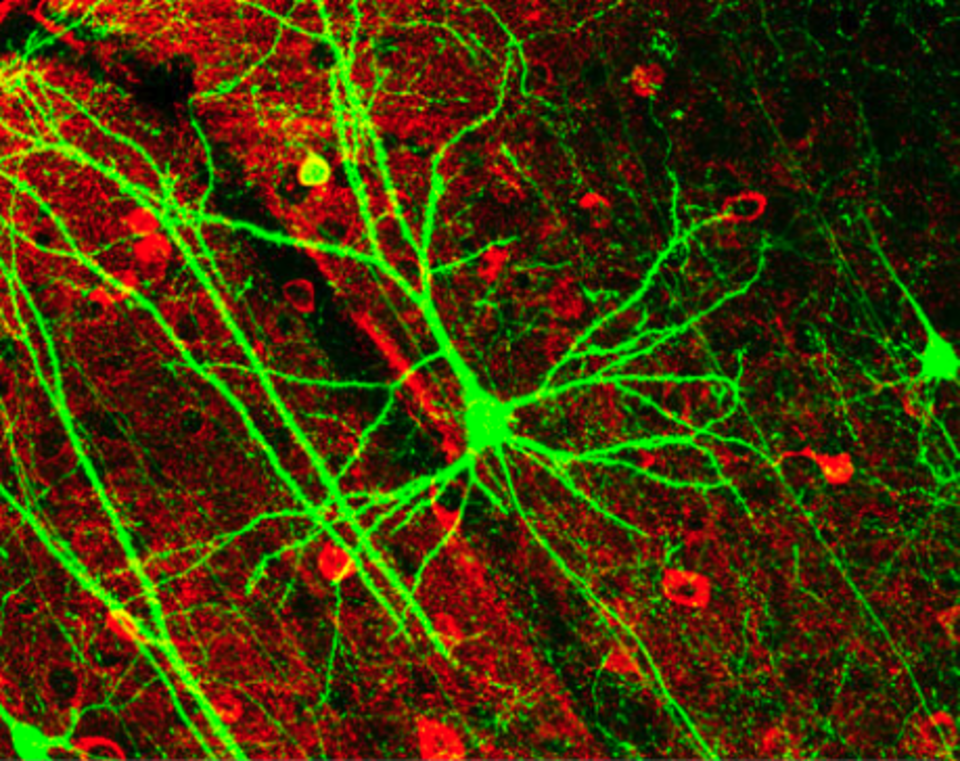
La plupart des neurones peuvent être caractérisés anatomiquement comme suit :
- Unipolaire : à un seul prolongement. Les cellules unipolaires sont exclusivement des neurones sensoriels. Leurs dendrites reçoivent l’information sensorielle, parfois directement du stimulus. Le corps cellulaire des neurones unipolaires se trouve toujours dans un ganglion. La réception sensorielle étant une fonction périphérique, le corps cellulaire est situé en périphérie, bien que plus proche du SNC au sein d’un ganglion. L’axone, issu des terminaisons dendritiques, passe devant le corps cellulaire dans le ganglion et pénètre dans le système nerveux central.
- Bipolaires : 1 axone et 1 dendrite. On les trouve principalement dans l’ épithélium olfactif et dans la rétine.
- Multipolaire : 1 axone et 2 dendrites ou plus
- Golgi I : neurones à longs prolongements axonaux ; exemples : cellules pyramidales, cellules de Purkinje et cellules de la corne antérieure
- Appareil de Golgi II : neurones dont le prolongement axonal se projette localement ; le meilleur exemple est la cellule granuleuse
- Anaxonique : où l'axone ne peut être distingué de la ou des dendrites.
- Pseudounipolaire : 1 processus qui sert alors à la fois d'axone et de dendrite
Autre
Certains types neuronaux uniques peuvent être identifiés selon leur localisation dans le système nerveux et leur forme particulière. En voici quelques exemples :Les cellules en panier sont des interneurones qui forment un plexus dense de terminaisons autour du soma des cellules cibles, présentes dans le cortex et le cervelet.
Classification fonctionnelle
Direction
- Les neurones afférents transmettent les informations des tissus et des organes au système nerveux central et sont également appelés neurones sensoriels .
- Les neurones efférents (neurones moteurs) transmettent les signaux du système nerveux central aux cellules effectrices.
- Les interneurones connectent les neurones au sein de régions spécifiques du système nerveux central.
Les termes afférents et efférents désignent également, de manière générale, les neurones qui, respectivement, apportent des informations au cerveau ou envoient des informations depuis celui-ci.
Action sur d'autres neurones
Un neurone influence d'autres neurones en libérant un neurotransmetteur qui se fixe à des récepteurs chimiques . L'effet sur le neurone postsynaptique est déterminé par le type de récepteur activé, et non par le neurone présynaptique ou le neurotransmetteur lui-même. Les récepteurs sont classés en trois grandes catégories : excitateurs (entraînant une augmentation de la fréquence de décharge), inhibiteurs (entraînant une diminution de la fréquence de décharge) et modulateurs (provoquant des effets durables non directement liés à la fréquence de décharge).le glutamate et le GABA , ont des actions globalement similaires. Le glutamate agit sur plusieurs types de récepteurs et exerce un effet excitateur sur les récepteurs ionotropes et un effet modulateur sur les récepteurs métabotropes . De même, le GABA agit sur plusieurs types de récepteurs, mais tous exercent un effet inhibiteur (chez l'animal adulte, du moins). Du fait de cette similarité, les neuroscientifiques désignent généralement les cellules libérant du glutamate comme des « neurones excitateurs » et celles libérant du GABA comme des « neurones inhibiteurs ». D'autres types de neurones présentent également des effets similaires, par exemple les motoneurones « excitateurs » de la moelle épinière qui libèrent de l'acétylcholine et les neurones spinaux « inhibiteurs » qui libèrent de la glycine .les cellules photoréceptrices de la rétine libèrent constamment le neurotransmetteur glutamate en l'absence de lumière. Les cellules bipolaires dites OFF sont, comme la plupart des neurones, excitées par le glutamate libéré. Cependant, les neurones cibles voisins, appelés cellules bipolaires ON, sont au contraire inhibés par le glutamate, car ils sont dépourvus de récepteurs ionotropes du glutamate typiques et expriment à la place une classe de récepteurs métabotropes inhibiteurs du glutamate. En présence de lumière, les photorécepteurs cessent de libérer du glutamate, ce qui lève l'inhibition des cellules bipolaires ON et les active ; simultanément, l'excitation des cellules bipolaires OFF est supprimée, ce qui les met au repos.la parvalbumine atténuent généralement le signal de sortie du neurone postsynaptique dans le cortex visuel , tandis que les neurones exprimant la somatostatine bloquent généralement les entrées dendritiques vers le neurone postsynaptique.
Modèles de décharge
Les neurones possèdent des propriétés électroréactives intrinsèques, telles que des profils d'oscillations transmembranaires intrinsèques . [ , les neurones peuvent être classés selon leurs caractéristiques électrophysiologiques :
- Activité tonique ou régulière. Certains neurones sont généralement constamment (toniquement) actifs, avec une fréquence de décharge constante. Exemple : les interneurones du neurostriatum .
- Phasique ou par salves . Les neurones qui déchargent par salves sont dits phasiques.
- Décharge rapide. Certains neurones sont remarquables par leur fréquence de décharge élevée, par exemple certains types d'interneurones inhibiteurs corticaux, les cellules du globus pallidus , les cellules ganglionnaires rétiniennes .
Neurotransmetteur
Cholinergique
L'acétylcholine est libérée par les neurones présynaptiques dans la fente synaptique. Elle agit comme ligand à la fois pour les canaux ioniques ligand-dépendants et pour les récepteurs muscariniques métabotropiques (RCPG) . Les récepteurs nicotiniques sont des canaux ioniques ligand-dépendants pentamériques composés de sous-unités alpha et bêta qui se lient à la nicotine . La liaison du ligand ouvre le canal, provoquant un influx d'ions Na +, une dépolarisation et augmentant la probabilité de libération présynaptique du neurotransmetteur. L'acétylcholine est synthétisée à partir de choline et d'acétyl-coenzyme A.
adrénergique
La noradrénaline est libérée par la plupart des neurones postganglionnaires du système nerveux sympathique sur deux types de récepteurs couplés aux protéines G (RCPG) : les récepteurs alpha-adrénergiques et les récepteurs bêta-adrénergiques . La noradrénaline est l’un des trois neurotransmetteurs catécholaminergiques les plus courants , et le plus abondant dans le système nerveux périphérique ; comme les autres catécholamines, elle est synthétisée à partir de la tyrosine .
GABAergique
Le GABA est l'un des deux neuroinhibiteurs du système nerveux central (SNC), avec la glycine. Il possède une fonction homologue à celle de l'acétylcholine (ACh) : il régule l'ouverture des canaux anioniques permettant aux ions Cl⁻ de pénétrer dans le neurone postsynaptique. Les ions Cl⁻ induisent une hyperpolarisation du neurone, diminuant ainsi la probabilité de déclenchement d'un potentiel d'action à mesure que le potentiel devient plus négatif (un seuil de potentiel positif doit être atteint pour qu'un potentiel d'action se déclenche). Le GABA est synthétisé à partir du glutamate par l'enzyme glutamate décarboxylase .
Glutamatergique
Le glutamate est l'un des deux principaux neurotransmetteurs excitateurs de la famille des acides aminés, avec l'aspartate . Les récepteurs du glutamate appartiennent à l'une des quatre catégories suivantes : trois sont des canaux ioniques ligand-dépendants et une est un récepteur couplé aux protéines G (souvent appelé RCPG).
- Les récepteurs AMPA et kaïnate fonctionnent comme des canaux cationiques perméables aux canaux cationiques Na + qui assurent une transmission synaptique excitatrice rapide.
Les récepteurs NMDA constituent un autre canal cationique plus perméable au Ca²⁺ . Leur fonctionnement dépend de la liaison du récepteur de la glycine, qui agit comme co- agoniste au sein du pore du canal. Les récepteurs NMDA sont inactifs en l'absence des deux ligands. Les récepteurs métabotropiques, ou RCPG, modulent la transmission synaptique et l'excitabilité postsynaptique.
- Le glutamate peut provoquer une excitotoxicité lorsque le flux sanguin cérébral est interrompu, entraînant des lésions cérébrales . En cas de diminution du flux sanguin, le glutamate est libéré par les neurones présynaptiques, ce qui provoque une activation plus importante des récepteurs NMDA et AMPA que la normale en dehors des situations de stress. Il en résulte une augmentation des concentrations de Ca²⁺ et de Na⁺ dans le neurone postsynaptique et des lésions cellulaires. Le glutamate est synthétisé à partir de l'acide aminé glutamine par l'enzyme glutamate synthase .
dopaminergique
La dopamine est un neurotransmetteur qui agit sur les récepteurs D1 (D1 et D5) couplés aux protéines Gs, ce qui augmente les taux d'AMPc et de PKA, et sur les récepteurs D2 (D2, D3 et D4), qui activent les récepteurs couplés aux protéines Gi et diminuent les taux d'AMPc et de PKA. La dopamine est impliquée dans la régulation de l'humeur et du comportement et module la neurotransmission pré- et post-synaptique. La perte de neurones dopaminergiques dans la substance noire est associée à la maladie de Parkinson . La dopamine est synthétisée à partir de l'acide aminé tyrosine . La tyrosine est convertie en lévodopa (ou L-DOPA ) par la tyrosine hydroxylase , puis la lévodopa en dopamine par la décarboxylase des acides aminés aromatiques .
sérotoninergique
La sérotonine (5-hydroxytryptamine, 5-HT) peut avoir une action excitatrice ou inhibitrice. Parmi ses quatre classes de récepteurs 5-HT, trois sont des récepteurs couplés aux protéines G (RCPG) et une est un canal cationique ligand-dépendant. La sérotonine est synthétisée à partir du tryptophane par la tryptophane hydroxylase , puis par la décarboxylase. Un déficit en 5-HT au niveau des neurones postsynaptiques est associé à la dépression . Des médicaments bloquant le transporteur présynaptique de la sérotonine , comme le Prozac et le Zoloft , sont utilisés pour traiter la dépression.
purinergique
L'ATP est un neurotransmetteur agissant à la fois sur les canaux ioniques ligand-dépendants ( récepteurs P2X ) et sur les récepteurs couplés aux protéines G (RCPG) ( récepteurs P2Y ). L'ATP est cependant surtout connu comme cotransmetteur . Cette signalisation purinergique peut également être médiée par d'autres purines comme l'adénosine , qui agit notamment sur les récepteurs P2Y.
Histaminergique
L'histamine est un neurotransmetteur et neuromodulateur monoaminergique . Les neurones histamino-producteurs se trouvent dans le noyau tubéromammillaire de l' hypothalamus . L'histamine intervient dans l'éveil et la régulation du cycle veille-sommeil.
Classification multimodale
Depuis 2012, la communauté des neurosciences cellulaires et computationnelles s'efforce d'établir une classification universelle des neurones, applicable à tous les neurones du cerveau et à toutes les espèces. Cette classification repose sur trois caractéristiques essentielles : l'électrophysiologie, la morphologie et le transcriptome individuel de chaque cellule. Outre son universalité, elle présente l'avantage de pouvoir également classer les astrocytes. L' Institut Allen pour la science du cerveau utilise largement une méthode appelée séquençage par patch-clamp, qui permet de mesurer simultanément ces trois caractéristiques . En 2023, un atlas cellulaire complet du cerveau humain adulte et en développement, aux niveaux transcriptionnel, épigénétique et fonctionnel, a été créé grâce à une collaboration internationale de chercheurs utilisant les approches de biologie moléculaire les plus pointues
Connectivité
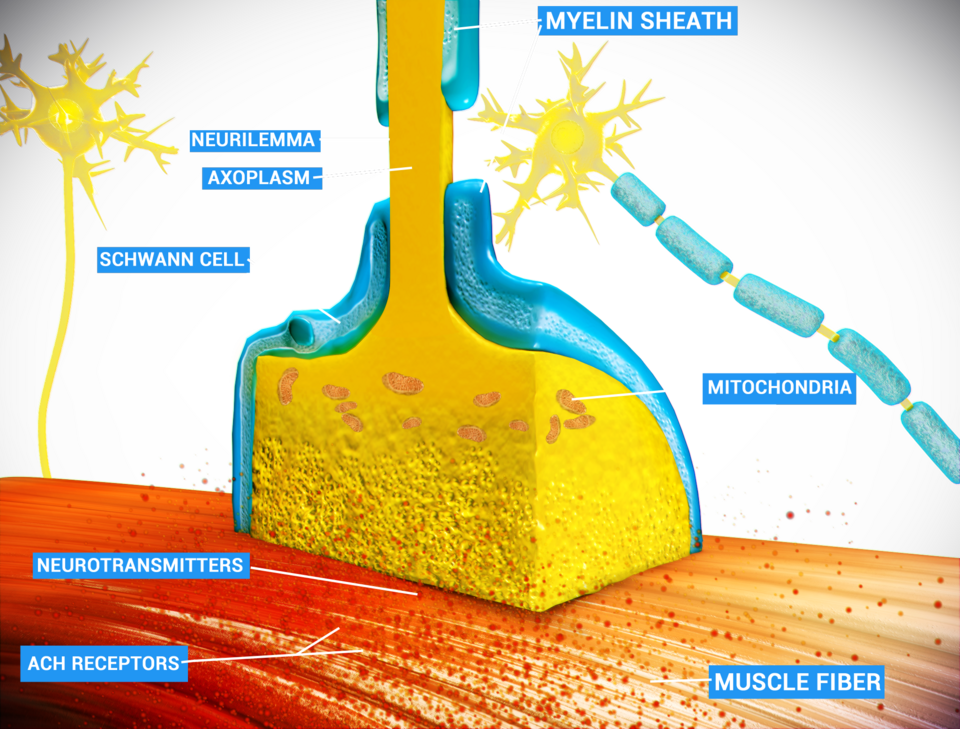
Les neurones communiquent entre eux par l'intermédiaire de synapses , où l' extrémité axonale d'une cellule entre en contact avec la dendrite, le soma ou, plus rarement, l'axone d'un autre neurone. Certains neurones, comme les cellules de Purkinje du cervelet, peuvent posséder plus de 1 000 ramifications dendritiques, établissant des connexions avec des dizaines de milliers d'autres cellules ; d'autres neurones, tels que les neurones magnocellulaires du noyau supraoptique , ne possèdent qu'une ou deux dendrites, chacune recevant des milliers de synapses.
Les synapses peuvent être excitatrices ou inhibitrices, augmentant ou diminuant respectivement l'activité du neurone cible. Certains neurones communiquent également via des synapses électriques, qui sont des jonctions directes et électriquement conductrices entre les cellules.
Lorsqu'un potentiel d'action atteint la terminaison axonale, il ouvre les canaux calciques voltage-dépendants , permettant aux ions calcium de pénétrer dans la terminaison. Le calcium provoque la fusion des vésicules synaptiques remplies de molécules de neurotransmetteurs avec la membrane, libérant ainsi leur contenu dans la fente synaptique. Les neurotransmetteurs diffusent à travers la fente synaptique et activent les récepteurs du neurone postsynaptique. L'augmentation de la concentration de calcium cytosolique dans la terminaison axonale déclenche la capture du calcium par les mitochondries, ce qui, à son tour, active le métabolisme énergétique mitochondrial pour produire de l'ATP et assurer la continuité de la neurotransmission.
Une autapse est une synapse dans laquelle l'axone d'un neurone se connecte à ses dendrites.
Le cerveau humain compte environ 8,6 × 10¹⁰ (quatre-vingt-six milliards) de neurones. Chaque neurone possède en moyenne 7 000 connexions synaptiques avec d’autres neurones. On estime que le cerveau d’un enfant de trois ans possède environ 10¹⁵ synapses (un quadrillion). Ce nombre diminue avec l’âge et se stabilise à l’âge adulte. Les estimations varient pour un adulte, allant de 10¹⁴ à 5 × 10¹⁴ synapses (100 à 500 billions).
Signalisation non électrochimique
Outre la signalisation électrique et chimique, des études suggèrent que les neurones du cerveau humain sain peuvent également communiquer par :
- force générée par l'élargissement des épines dendritiques
- le transfert de protéines – protéines transportées transneuronalement (TNTP)
Leur activité peut également être modulée par des facteurs environnementaux et des hormones sécrétées par d'autres parties de l'organisme , lesquelles peuvent être influencées plus ou moins directement par les neurones. Ceci s'applique également aux neurotrophines telles que le BDNF . Le microbiote intestinal est lui aussi connecté au cerveau . Les neurones communiquent également avec la microglie , principale cellule immunitaire du cerveau, via des sites de contact spécialisés appelés « jonctions somatiques ». Ces connexions permettent à la microglie de surveiller et de réguler en permanence les fonctions neuronales, et d'exercer une neuroprotection en cas de besoin
Mécanismes de propagation des potentiels d'action
La membrane cellulaire de l'axone et du soma contient des canaux ioniques voltage-dépendants qui permettent au neurone de générer et de propager un signal électrique (un potentiel d'action). Certains neurones génèrent également des oscillations du potentiel membranaire en dessous du seuil. Ces signaux sont générés et propagés par des ions porteurs de charge, notamment le sodium (Na + ), le potassium (K + ), le chlorure (Cl- ) et le calcium (Ca2 + ) .
Plusieurs stimuli peuvent activer un neurone et induire une activité électrique, notamment la pression , l'étirement, les neurotransmetteurs et les variations du potentiel électrique transmembranaire. Ces stimuli provoquent l'ouverture de canaux ioniques spécifiques au sein de la membrane cellulaire, entraînant un flux d'ions et modifiant ainsi le potentiel membranaire. Les neurones doivent conserver les propriétés électriques spécifiques qui définissent leur type.
Les neurones et axones fins nécessitent moins d'énergie pour produire et propager les potentiels d'action, tandis que les axones plus épais transmettent les influx nerveux plus rapidement. Afin de minimiser cette énergie tout en maintenant une conduction rapide, de nombreux neurones possèdent des gaines isolantes de myéline autour de leurs axones. Ces gaines sont formées par les cellules gliales : les oligodendrocytes dans le système nerveux central et les cellules de Schwann dans le système nerveux périphérique. La gaine permet aux potentiels d'action de se propager plus rapidement que dans les axones non myélinisés de même diamètre, tout en consommant moins d'énergie. Dans les nerfs périphériques, la gaine de myéline s'étend normalement le long de l'axone par segments d'environ 1 mm de long, ponctués de nœuds de Ranvier non myélinisés , qui contiennent une forte densité de canaux ioniques voltage-dépendants. La sclérose en plaques est une maladie neurologique résultant de la démyélinisation des axones dans le système nerveux central.
Certains neurones ne génèrent pas de potentiels d'action, mais plutôt un signal électrique gradué , qui induit une libération graduelle de neurotransmetteurs. Ces neurones non-potentiels d'action sont généralement des neurones sensoriels ou des interneurones, car ils ne peuvent pas transmettre de signaux sur de longues distances.
Codage neuronal
Le codage neuronal s'intéresse à la manière dont les neurones représentent les informations sensorielles et autres dans le cerveau. L'objectif principal de l'étude du codage neuronal est de caractériser la relation entre le stimulus et les réponses neuronales individuelles ou collectives , ainsi que les relations entre les activités électriques des neurones au sein de cet ensemble. On pense que les neurones peuvent coder des informations à la fois numériques et analogiques .
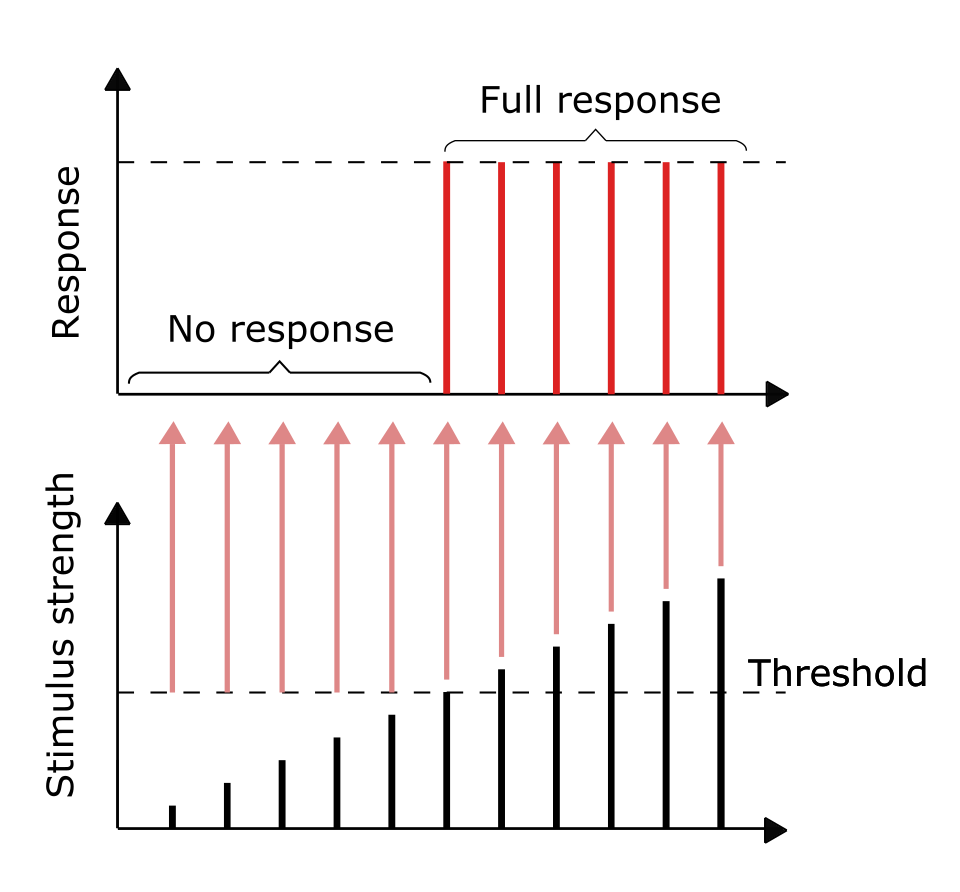
Principe du tout ou rien
Parmi les autres types de récepteurs, on trouve les récepteurs à adaptation rapide ou phasiques, dont l'activité diminue ou s'arrête sous l'effet d'une stimulation constante. C'est le cas, par exemple, de la peau : au toucher, les neurones s'activent, mais si la pression exercée reste constante, l'activité neuronale cesse. Les neurones de la peau et des muscles sensibles à la pression et aux vibrations possèdent des structures accessoires de filtrage qui optimisent leur fonctionnement.
Le corpuscule de Pacini est une de ces structures. Il est constitué de couches concentriques, semblables à celles d'un oignon, qui se forment autour de la terminaison axonale. Lorsqu'une pression est appliquée et que le corpuscule se déforme, un stimulus mécanique est transmis à l'axone, déclenchant un potentiel d'action. Si la pression est stable, le stimulus cesse ; ainsi, ces neurones répondent généralement par une dépolarisation transitoire lors de la déformation initiale, puis à nouveau lorsque la pression est relâchée, ce qui provoque une nouvelle modification de la forme du corpuscule. D'autres types d'adaptation sont importants pour étendre la fonction de plusieurs autres neurones.
Bien que l'on ait longtemps supposé que les neurones ne produisent qu'une réponse maximale stéréotypée ou aucune réponse du tout, des recherches récentes suggèrent que cette hypothèse est partiellement erronée. En effet, si les neurones génèrent ou non un potentiel d'action, l'amplitude et la durée de ce potentiel peuvent varier considérablement, permettant ainsi au neurone d'encoder des informations, au moins par l'intensité du potentiel d'action. De plus, il a été démontré que l'information analogique véhiculée par le potentiel d'action peut survivre et se propager sur des distances que l'on croyait auparavant impossibles. Ce mécanisme est considéré comme une méthode d'encodage de l'information très efficace, comparée aux théories classiques de codage temporel et de fréquence, et permettrait de transférer environ quatre fois plus d'informations que ce que l'on pensait jusqu'alors.
Étymologie et orthographe
L'anatomiste allemand Heinrich Wilhelm Waldeyer a introduit le terme neurone en 1891, basé sur le grec ancien νεῦρον neuron « tendon, cordon, nerf ».
Le mot a été adopté en français sous l'orthographe « neurone » . Cette orthographe a également été utilisée par de nombreux auteurs en anglais, mais elle est maintenant devenue rare en anglais américain et peu courante en anglais britannique.
Certains travaux antérieurs ont utilisé la cellule nerveuse ( cellule nervose ), comme adopté dans l'article de Camillo Golgi de 1873 sur la découverte de la technique de coloration à l'argent utilisée pour visualiser le tissu nerveux sous microscopie optique.
Histoire
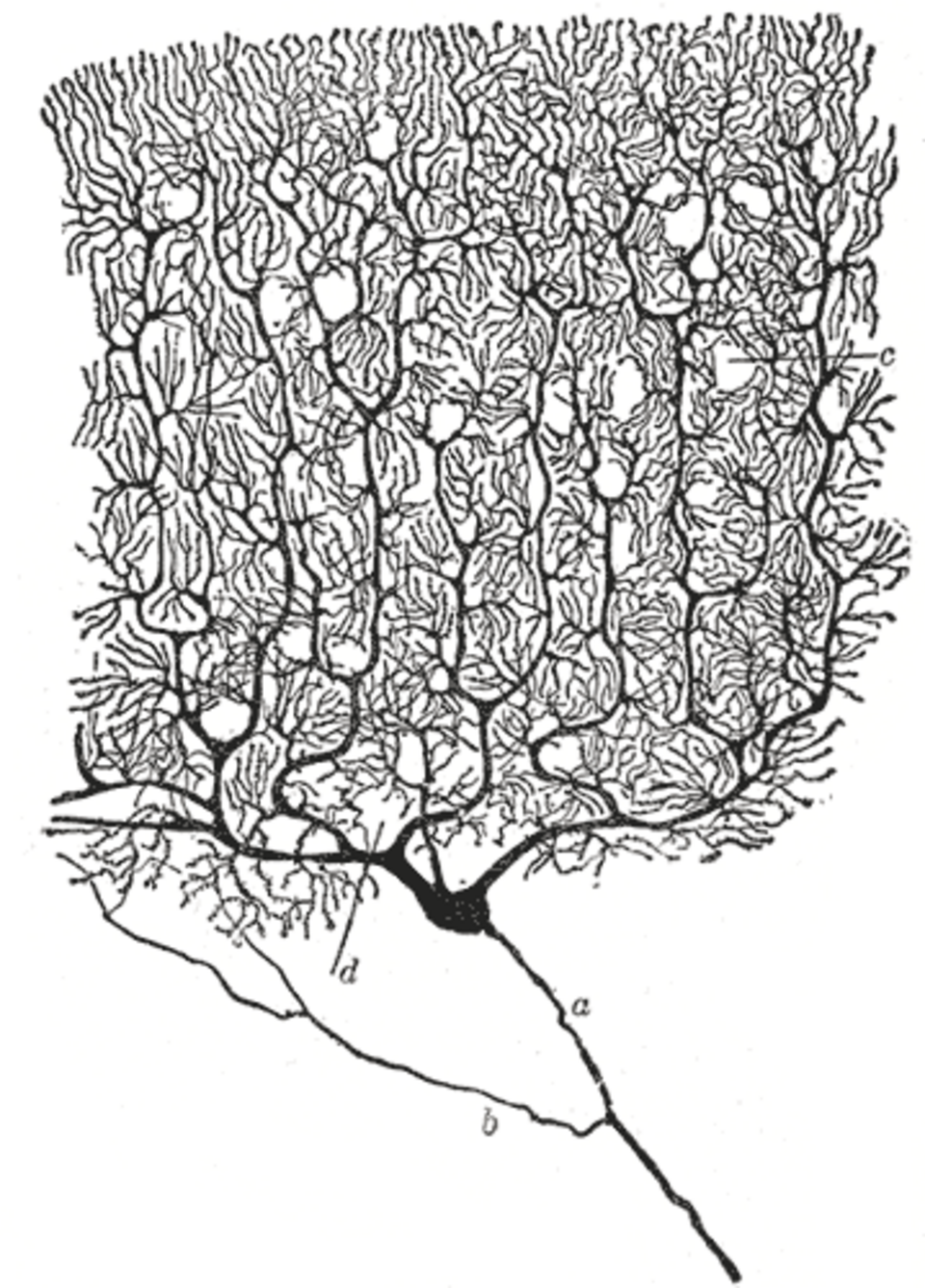
La place du neurone en tant qu'unité fonctionnelle primaire du système nerveux a été reconnue pour la première fois à la fin du XIXe siècle grâce aux travaux de l'anatomiste espagnol Santiago Ramón y Cajal .
Pour rendre visible la structure des neurones individuels, Ramón y Cajal a amélioré un procédé de coloration à l'argent qui avait été mis au point par Camillo Golgi . Le procédé amélioré implique une technique appelée « double imprégnation » et est toujours utilisé.
En 1888, Ramón y Cajal publia un article sur le cervelet des oiseaux. Dans cet article, il affirmait ne pas avoir trouvé de preuves d' anastomose entre les axones et les dendrites et qualifiait chaque élément nerveux de « canton autonome ». Cette conception devint la doctrine du neurone , l'un des principes fondamentaux des neurosciences modernes .
En 1891, l'anatomiste allemand Heinrich Wilhelm Waldeyer a écrit une revue très influente de la doctrine du neurone dans laquelle il a introduit le terme neurone pour décrire l'unité anatomique et physiologique du système nerveux.
Les colorations par imprégnation argentique sont une méthode utile pour les recherches neuroanatomiques car, pour des raisons inconnues, elles ne colorent qu'un faible pourcentage de cellules dans un tissu, exposant la microstructure complète des neurones individuels sans beaucoup de chevauchement avec d'autres cellules.
doctrine neuronale
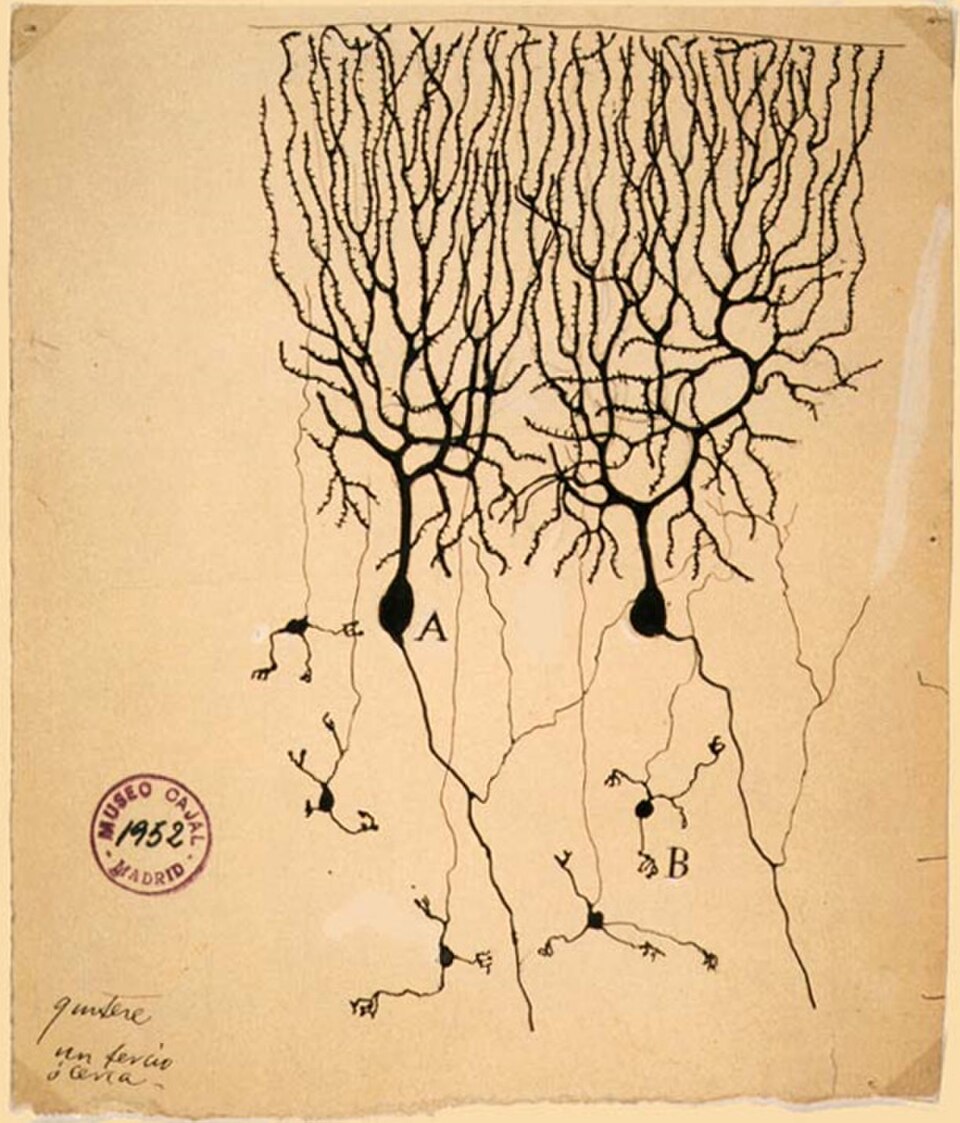
La théorie du neurone, aujourd'hui fondamentale, postule que les neurones constituent les unités structurales et fonctionnelles de base du système nerveux. Élaborée par Santiago Ramón y Cajal à la fin du XIXe siècle, elle considère les neurones comme des cellules distinctes (non interconnectées en un réseau), fonctionnant comme des unités métaboliquement indépendantes.
Des découvertes ultérieures ont permis d'affiner cette doctrine. Par exemple, les cellules gliales , qui ne sont pas neuronales, jouent un rôle essentiel dans le traitement de l'information. De plus, les synapses électriques sont plus fréquentes qu'on ne le pensait auparavant, et consistent en des connexions cytoplasmiques directes entre les neurones ; en fait, les neurones peuvent former des couplages encore plus étroits : l'axone géant du calmar résulte de la fusion de plusieurs axones.
Ramón y Cajal a également postulé la loi de polarisation dynamique, selon laquelle un neurone reçoit des signaux au niveau de ses dendrites et de son corps cellulaire et les transmet, sous forme de potentiels d'action, le long de l'axone dans une seule direction : en s'éloignant du corps cellulaire. La loi de polarisation dynamique présente d'importantes exceptions ; les dendrites peuvent servir de sites de sortie synaptique des neurones et les axones peuvent recevoir des entrées synaptiques.
Modélisation compartimentale des neurones
Bien que les neurones soient souvent décrits comme les « unités fondamentales » du cerveau, ils effectuent des calculs internes. Les neurones intègrent les informations au sein de leurs dendrites, et cette complexité est occultée par les modèles qui les considèrent comme une unité fondamentale. Les branches dendritiques peuvent être modélisées comme des compartiments spatiaux, dont l’activité est liée aux propriétés passives de la membrane, mais peut également varier en fonction des entrées synaptiques. La modélisation compartimentale des dendrites est particulièrement utile pour comprendre le comportement des neurones trop petits pour être enregistrés par électrodes, comme c’est le cas chez Drosophila melanogaster .
Neurones du cerveau
Le nombre de neurones dans le cerveau varie considérablement d'une espèce à l'autre. Chez l'humain, on estime à 10–20 milliards le nombre de neurones dans le cortex cérébral et à 55–70 milliards celui du cervelet . À l'inverse, le nématode Caenorhabditis elegans ne possède que 302 neurones, ce qui en fait un organisme modèle idéal , les scientifiques ayant pu cartographier l'ensemble de son réseau neuronal. La drosophile (Drosophila melanogaster) , un sujet fréquemment utilisé en biologie, possède environ 100 000 neurones et présente de nombreux comportements complexes. De nombreuses propriétés des neurones, du type de neurotransmetteurs utilisés à la composition des canaux ioniques, sont conservées d'une espèce à l'autre, permettant ainsi aux scientifiques d'étudier des processus se déroulant chez des organismes plus complexes dans des systèmes expérimentaux beaucoup plus simples.
Troubles neurologiques
La maladie d’Alzheimer (MA), également appelée simplement Alzheimer , est une maladie neurodégénérative caractérisée par un déclin cognitif progressif , associé à une diminution des capacités fonctionnelles au quotidien et à des symptômes neuropsychiatriques ou des changements comportementaux. Le symptôme précoce le plus frappant est la perte de mémoire à court terme ( amnésie ), qui se manifeste généralement par de légers oublis s’accentuant progressivement avec l’évolution de la maladie, tandis que les souvenirs anciens sont relativement préservés. À mesure que la maladie progresse, les troubles cognitifs (intellectuels) s’étendent aux domaines du langage ( aphasie ), de la motricité fine ( apraxie ) et de la reconnaissance ( agnosie ), et des fonctions telles que la prise de décision et la planification sont altérées.
La maladie de Parkinson (MP), également appelée sclérose latérale amyotrophique ( SLA ), est une affection neurodégénérative du système nerveux central qui altère souvent la motricité et la parole. Elle fait partie des troubles du mouvement . Elle se caractérise par une rigidité musculaire, des tremblements , un ralentissement des mouvements ( bradykinésie ) et, dans les cas les plus graves, une perte de mobilité ( akinésie ). Les symptômes principaux résultent d'une diminution de la stimulation du cortex moteur par les noyaux gris centraux , généralement due à une production et une action insuffisantes de dopamine, neurotransmetteur produit par les neurones dopaminergiques du cerveau. Les symptômes secondaires peuvent inclure des troubles cognitifs importants et des troubles du langage plus subtils. La MP est une maladie chronique et progressive.
La myasthénie est une maladie neuromusculaire qui provoque une faiblesse musculaire fluctuante et une fatigabilité même lors d'activités simples. Cette faiblesse est généralement due à des anticorps circulants qui bloquent les récepteurs de l'acétylcholine au niveau de la jonction neuromusculaire postsynaptique, inhibant ainsi l'effet stimulant de ce neurotransmetteur. Le traitement de la myasthénie repose sur l'administration d'immunosuppresseurs , d'inhibiteurs de la cholinestérase et, dans certains cas, sur la thymectomie .
Démyélinisation
La démyélinisation est un processus caractérisé par la perte progressive de la gaine de myéline qui enveloppe les fibres nerveuses. Lorsque la myéline se détériore, la conduction des signaux nerveux peut être fortement altérée, voire interrompue, et le nerf finit par s'atrophier. La démyélinisation peut affecter le système nerveux central et périphérique, contribuant à diverses affections neurologiques telles que la sclérose en plaques , le syndrome de Guillain-Barré et la polyneuropathie inflammatoire démyélinisante chronique . Bien que la démyélinisation soit souvent causée par une réaction auto-immune , elle peut également être provoquée par des infections virales, des troubles métaboliques, des traumatismes et certains médicaments.
Dégénérescence axonale
Bien que la plupart des réponses aux lésions comprennent une signalisation d'influx de calcium favorisant la cicatrisation des parties sectionnées, les lésions axonales entraînent initialement une dégénérescence axonale aiguë , caractérisée par la séparation rapide des extrémités proximale et distale, survenant dans les 30 minutes suivant la lésion . La dégénérescence s'accompagne d'un gonflement de l' axolemme , aboutissant à une formation de granules. Une désintégration granulaire du cytosquelette axonal et des organites internes se produit après la dégradation de l'axolemme. Parmi les changements précoces, on observe une accumulation de mitochondries dans les régions paranodales au site de la lésion. Le réticulum endoplasmique se dégrade, les mitochondries gonflent et finissent par se désintégrer. Cette désintégration dépend des protéases ubiquitine et calpaïne (provoquées par l'influx d'ions calcium), suggérant que la dégénérescence axonale est un processus actif aboutissant à une fragmentation complète. Ce processus dure environ 24 heures dans le SNP et plus longtemps dans le SNC. Les voies de signalisation conduisant à la dégénérescence de l'axolemme sont inconnues.
Développement
Chez l'embryon, les neurones se développent initialement à partir du tube neural . Ce dernier comporte trois couches : la zone ventriculaire , la zone intermédiaire et la zone marginale. La zone ventriculaire entoure le canal central du tube et se transforme en épendyme . La division cellulaire de la zone ventriculaire forme la zone intermédiaire, qui s'étend jusqu'à la couche la plus externe du tube neural, appelée la pie-mère. La substance grise du cerveau provient de la zone intermédiaire. Les prolongements neuronaux de cette zone, une fois myélinisés , constituent la zone marginale et forment la substance blanche du cerveau .
La différenciation des neurones est ordonnée selon leur taille. Les grands neurones moteurs sont les premiers. Les plus petits neurones sensoriels, ainsi que les cellules gliales, se différencient à la naissance .
La neurogenèse adulte est possible, et des études sur l'âge des neurones humains suggèrent que ce processus ne concerne qu'une minorité de cellules et que la grande majorité des neurones du néocortex se forment avant la naissance et persistent sans être remplacés. L'étendue de la neurogenèse adulte chez l'humain et sa contribution à la cognition font l'objet de controverses, comme en témoignent des résultats contradictoires publiés en 2018.
L’organisme contient différents types de cellules souches capables de se différencier en neurones. Des chercheurs ont découvert un moyen de transformer des cellules de la peau humaine en cellules nerveuses grâce à la transdifférenciation , un processus dans lequel « les cellules sont forcées d’adopter de nouvelles identités ».
Au cours de la neurogenèse dans le cerveau des mammifères, les cellules progénitrices et souches passent de divisions prolifératives à des divisions différenciées. Cette progression conduit aux neurones et aux cellules gliales qui peuplent les couches corticales. Les modifications épigénétiques jouent un rôle clé dans la régulation de l'expression des gènes dans les cellules souches neurales en différenciation et sont essentielles à la détermination du destin cellulaire dans le cerveau des mammifères, tant au cours du développement qu'à l'âge adulte. Ces modifications épigénétiques comprennent la méthylation de la cytosine de l'ADN en 5-méthylcytosine et la déméthylation de la 5-méthylcytosine . La méthylation de la cytosine de l'ADN est catalysée par les ADN méthyltransférases (DNMT) . La déméthylation de la 5-méthylcytosine est catalysée en plusieurs étapes par les enzymes TET , qui réalisent des réactions d'oxydation (par exemple, la conversion de la 5-méthylcytosine en 5-hydroxyméthylcytosine ), et par les enzymes de la voie de réparation par excision de base (BER) de l'ADN
Au cours des différentes étapes du développement du système nerveux des mammifères, deux processus de réparation de l'ADN sont utilisés pour réparer les cassures double brin. Ces voies sont la réparation par recombinaison homologue, utilisée dans les cellules précurseurs neurales en prolifération , et la jonction d'extrémités non homologues, utilisée principalement aux stades de développement ultérieurs
La communication intercellulaire entre les neurones en développement et la microglie est également indispensable à une neurogenèse et à un développement cérébral corrects.