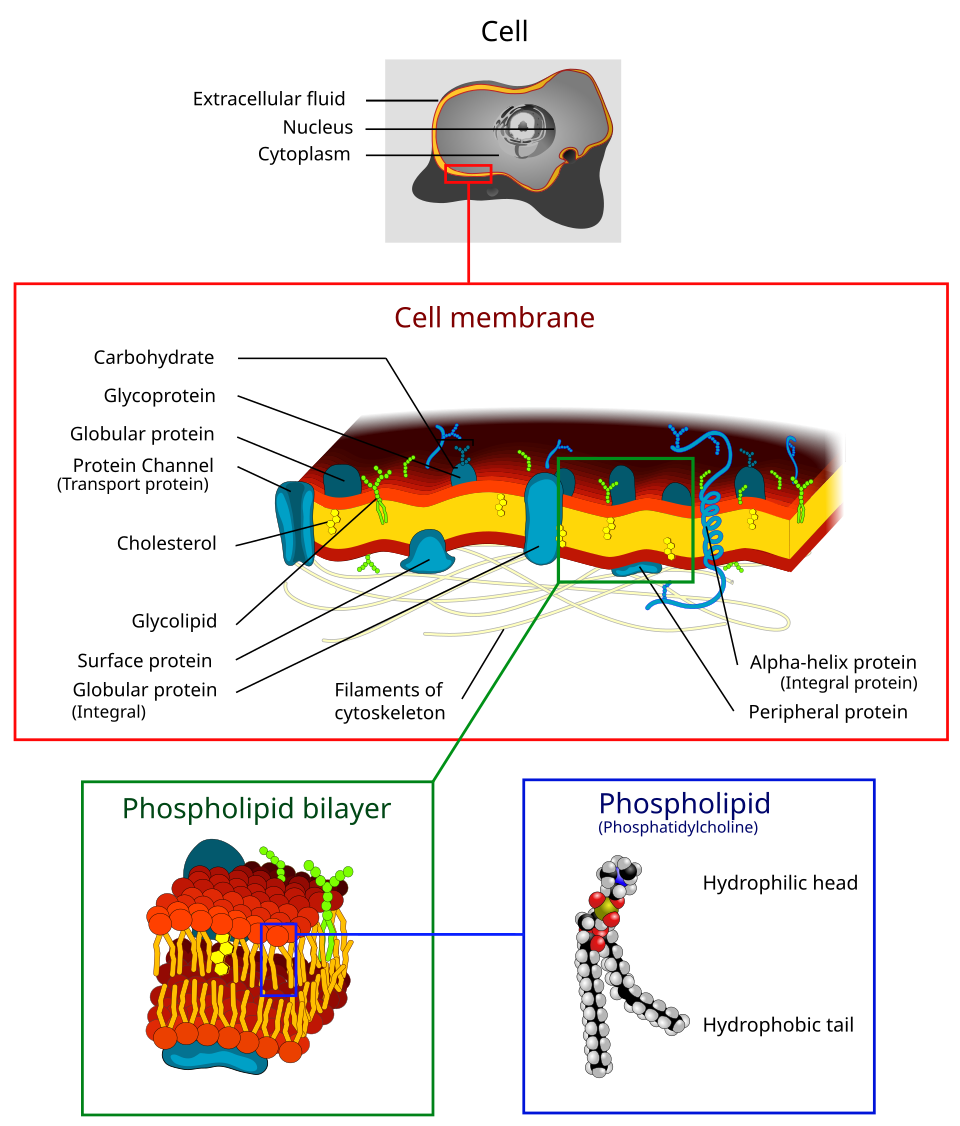
La membrane cellulaire (également appelée membrane plasmique ou membrane cytoplasmique , et historiquement désignée sous le nom de plasmalemme ) est une membrane biologique semi-perméable qui sépare et protège l' intérieur de la cellule du milieu extérieur (l'espace extracellulaire). La membrane cellulaire est une bicouche lipidique , généralement composée de phospholipides et de glycolipides ; chez les eucaryotes et certaines archées , on trouve également des stérols (comme le cholestérol chez les animaux) intercalés entre ces deux couches, ce qui permet de maintenir une fluidité membranaire appropriée à différentes températures. La membrane contient aussi des protéines membranaires , notamment des protéines intégrales qui traversent la membrane et servent de transporteurs , et des protéines périphériques qui se fixent à la surface de la membrane cellulaire et agissent comme des enzymes pour faciliter les interactions avec l'environnement cellulaire. Les glycolipides inclus dans la couche lipidique externe ont une fonction similaire.
La membrane cellulaire contrôle les échanges de substances entre l'intérieur et l'extérieur de la cellule, étant sélectivement perméable aux ions et aux molécules organiques. De plus, les membranes cellulaires interviennent dans divers processus cellulaires tels que l'adhésion cellulaire , la conductivité ionique et la signalisation cellulaire . Elles servent également de surface d'attachement à plusieurs structures extracellulaires, notamment la paroi cellulaire et le glycocalyx (enveloppe glucidique ) , ainsi qu'au cytosquelette (réseau intracellulaire de fibres protéiques ). En biologie synthétique, les membranes cellulaires peuvent être reconstituées artificiellement .
La découverte des cellules par Robert Hooke en 1665 a conduit à la proposition de la théorie cellulaire . Initialement, on pensait que toutes les cellules possédaient une paroi cellulaire rigide, car seules les cellules végétales étaient observables à l'époque. Les microscopistes se sont concentrés sur la paroi cellulaire pendant plus de 150 ans, jusqu'aux progrès de la microscopie. Au début du XIXe siècle, après la découverte de la possibilité de séparer les cellules végétales, on a reconnu que les cellules étaient des entités distinctes, non connectées entre elles et délimitées par des parois cellulaires individuelles. Cette théorie a été étendue aux cellules animales, suggérant un mécanisme universel de protection et de développement cellulaire.Dans la seconde moitié du XIXe siècle, la microscopie n'était pas encore suffisamment avancée pour distinguer les membranes cellulaires des parois cellulaires. Cependant, certains microscopistes ont correctement identifié à cette époque que, bien qu'invisibles, les membranes cellulaires existaient dans les cellules animales, comme en témoignait le mouvement intracellulaire de composants (à l'intérieur mais pas à l'extérieur de la cellule), et que ces membranes n'étaient pas l'équivalent d'une paroi cellulaire végétale . On a également déduit que les membranes cellulaires n'étaient pas des éléments vitaux pour toutes les cellules. Nombreux étaient ceux qui réfutaient encore l'existence des membranes cellulaires jusqu'à la fin du XIXe siècle. En 1890, une révision de la théorie cellulaire affirmait que les membranes cellulaires existaient, mais qu'elles n'étaient que des structures secondaires. Ce n'est qu'avec des études ultérieures sur l'osmose et la perméabilité que les membranes cellulaires ont été davantage reconnues. En 1895, Ernest Overton a proposé que les membranes cellulaires étaient composées de lipides.
L'hypothèse de la bicouche lipidique, proposée en 1925 par Gorter et Grendel a suscité des interrogations quant à la description de la structure bicouche de la membrane cellulaire, en se basant sur des études cristallographiques et des observations de bulles de savon. Afin de confirmer ou d'infirmer cette hypothèse, des chercheurs ont mesuré l'épaisseur de la membrane. Ils ont extrait les lipides de globules rouges humains et mesuré la surface qu'ils recouvraient une fois étalés sur l'eau. Les globules rouges matures de mammifères étant dépourvus de noyau et d'organites cytoplasmiques, la membrane plasmique est la seule structure contenant des lipides dans la cellule. Par conséquent, on peut supposer que tous les lipides extraits des cellules se trouvaient dans leurs membranes plasmiques. Le rapport entre la surface d'eau recouverte par les lipides extraits et la surface calculée pour les globules rouges dont ils proviennent était d'environ 2:1, ce qui a permis de conclure que la membrane plasmique contient une bicouche lipidique
En 1925, Fricke détermina que l'épaisseur des membranes des érythrocytes et des levures se situait entre 3,3 et 4 nm, une épaisseur compatible avec une monocouche lipidique. Le choix de la constante diélectrique utilisée dans ces études fut remis en question, mais des tests ultérieurs ne permirent pas d'infirmer les résultats de l'expérience initiale. Parallèlement, le leptoscope fut inventé pour mesurer les membranes très fines en comparant l'intensité de la lumière réfléchie par un échantillon à celle d'une membrane de référence d'épaisseur connue. Cet instrument permettait de mesurer des épaisseurs dépendant du pH et de la présence de protéines membranaires, allant de 8,6 à 23,2 nm, les valeurs les plus faibles confirmant l'hypothèse de la bicouche lipidique. Plus tard, dans les années 1930, le modèle de structure membranaire généralement admis fut le modèle paucimoléculaire de Davson et Danielli (1935). Ce modèle était basé sur des études de la tension superficielle entre les huiles et les œufs d'échinodermes . Étant donné que les valeurs de tension superficielle semblaient bien inférieures à celles attendues pour une interface huile-eau, on a supposé qu'une substance était responsable de la diminution des tensions interfaciales à la surface des cellules. Il a été suggéré qu'une bicouche lipidique se trouvait entre deux fines couches protéiques. Le modèle paucimoléculaire a immédiatement connu un grand succès et a dominé les études sur les membranes cellulaires pendant les 30 années suivantes, jusqu'à ce qu'il soit concurrencé par le modèle de la mosaïque fluide de Singer et Nicolson (1972).
Malgré les nombreux modèles de membrane cellulaire proposés avant le modèle de la mosaïque fluide , ce dernier demeure l'archétype principal de la membrane cellulaire, longtemps après son apparition dans les années 1970. Bien que le modèle de la mosaïque fluide ait été modernisé pour intégrer les découvertes contemporaines, ses principes fondamentaux restent inchangés : la membrane est une bicouche lipidique composée de têtes hydrophiles externes et d'un intérieur hydrophobe où les protéines peuvent interagir avec les têtes hydrophiles par des interactions polaires. Cependant, les protéines qui traversent totalement ou partiellement la bicouche possèdent des acides aminés hydrophobes qui interagissent avec l'intérieur lipidique non polaire. Le modèle de la mosaïque fluide a non seulement fourni une représentation précise de la mécanique membranaire, mais il a également permis d'approfondir l'étude des forces hydrophobes, qui deviendront par la suite une limitation descriptive essentielle pour la description des macromolécules biologiques .
Pendant des siècles, les scientifiques cités ont divergé quant à l'importance de la structure qu'ils identifiaient comme la membrane cellulaire. Pendant près de deux siècles, les membranes ont été observées, mais leur rôle essentiel dans la fonction cellulaire a été largement négligé. Ce n'est qu'au XXe siècle que l'importance de la membrane cellulaire a été reconnue. Finalement, deux scientifiques, Gorter et Grendel (1925), ont découvert que la membrane était composée de lipides. À partir de là, ils ont émis l'hypothèse que cette structure devait s'organiser en couches. Des études plus approfondies ont permis d'établir, en comparant la somme des surfaces cellulaires et des surfaces lipidiques, un rapport de 2:1, jetant ainsi les bases de la structure bicouche connue aujourd'hui. Cette découverte a initié de nombreuses études à travers le monde et dans divers domaines scientifiques, confirmant ainsi la large acceptation de la structure et des fonctions de la membrane cellulaire.
Cette structure a été désignée de diverses manières par différents auteurs : ectoplaste ( de Vries , 1885) , plasmahaut (peau plasmique, Pfeffer , 1877, 1891) , hautschicht (couche cutanée, Pfeffer, 1886 ; terme employé avec une signification différente par Hofmeister , 1867), membrane plasmique (Pfeffer, 1900) , membrane plasmique, membrane cytoplasmique, enveloppe cellulaire et membrane cellulaire . Certains auteurs, ne croyant pas à l’existence d’une frontière fonctionnelle perméable à la surface de la cellule, ont préféré utiliser le terme plasmalemme (inventé par Mast, 1924) pour désigner la région externe de la cellule
Composition
Les membranes cellulaires contiennent diverses molécules biologiques , notamment des lipides et des protéines. Leur composition n'est pas fixe ; elle évolue constamment pour assurer la fluidité membranaire et s'adapter aux variations de l'environnement, et fluctue même au cours des différentes étapes du développement cellulaire. Plus précisément, la quantité de cholestérol dans la membrane des neurones primaires humains varie, et cette variation de composition influence la fluidité membranaire tout au long du développement.
L'incorporation ou l'élimination de matière dans la membrane se fait par divers mécanismes :
- La fusion des vésicules intracellulaires avec la membrane ( exocytose ) permet non seulement d'excréter le contenu de la vésicule, mais aussi d'incorporer les composants de sa membrane à la membrane cellulaire. La membrane peut former des protrusions autour de la substance extracellulaire qui se détachent pour former des vésicules ( endocytose ).
- Si une membrane est en continuité avec une structure tubulaire constituée du matériau de la membrane, alors le matériau provenant du tube peut être aspiré en continu dans la membrane.
- Bien que la concentration des composants membranaires dans la phase aqueuse soit faible (les composants membranaires stables ont une faible solubilité dans l'eau), il existe un échange de molécules entre les phases lipidique et aqueuse.
Lipides
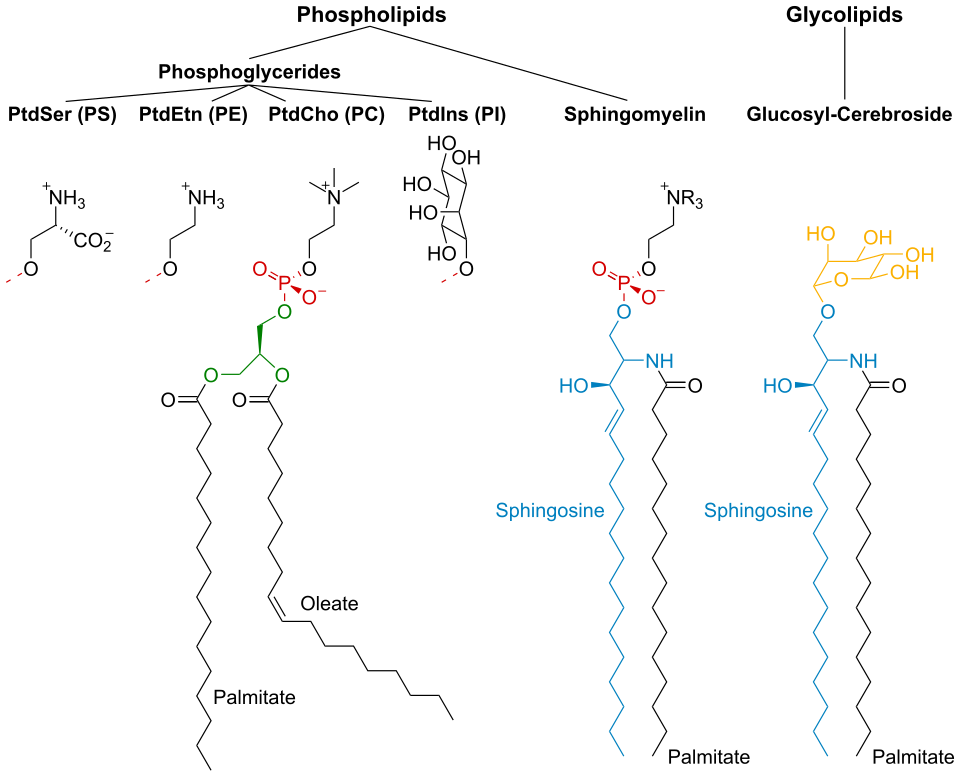
La membrane cellulaire est constituée de trois classes de lipides amphipathiques : les phospholipides , les glycolipides et les stérols . Leur proportion varie selon le type cellulaire, mais dans la plupart des cas, les phospholipides sont les plus abondants, représentant souvent plus de 50 % des lipides membranaires. Les glycolipides ne représentent qu’une infime partie (environ 2 %) et les stérols constituent le reste. Dans les globules rouges , la membrane plasmique est composée à 30 % de lipides. Cependant, chez la majorité des cellules eucaryotes, la composition des membranes plasmiques est d’environ moitié lipides et moitié protéines (en poids).
Les chaînes grasses des phospholipides et des glycolipides contiennent généralement un nombre pair d'atomes de carbone, typiquement entre 16 et 20. Les acides gras à 16 et 18 atomes de carbone sont les plus courants. Les acides gras peuvent être saturés ou insaturés, la configuration des doubles liaisons étant presque toujours « cis ». La longueur et le degré d'insaturation des chaînes d'acides gras ont un impact important sur la fluidité membranaire : les lipides insaturés créent une courbure qui empêche les acides gras de s'agencer de manière aussi compacte, abaissant ainsi la température de fusion (et augmentant la fluidité) de la membrane. La capacité de certains organismes à réguler la fluidité de leurs membranes cellulaires en modifiant leur composition lipidique est appelée adaptation homéovisqueuse .
La membrane entière est maintenue par des interactions non covalentes des queues hydrophobes ; cependant, sa structure est relativement fluide et non rigide. Dans des conditions physiologiques, les molécules de phospholipides de la membrane cellulaire sont à l’ état de cristal liquide . Cela signifie que les molécules lipidiques peuvent diffuser librement et présentent une diffusion latérale rapide le long de la bicouche dans laquelle elles se trouvent . Toutefois, l’échange de molécules de phospholipides entre les feuillets intracellulaire et extracellulaire de la bicouche est un processus très lent. Les radeaux lipidiques et les cavéoles sont des exemples de microdomaines enrichis en cholestérol dans la membrane cellulaire . De plus, une fraction des lipides en contact direct avec les protéines membranaires intégrales, et fortement liée à la surface de la protéine, est appelée enveloppe lipidique annulaire ; elle se comporte comme une partie du complexe protéique.
Le cholestérol est normalement présent en quantités variables dans les membranes cellulaires, dans les espaces irréguliers entre les chaînes hydrophobes des lipides membranaires, où il confère à la membrane une rigidité et un renforcement. De plus, la quantité de cholestérol dans les membranes biologiques varie selon les organismes, les types cellulaires, et même au sein d'une même cellule. Composant majeur des membranes plasmiques, le cholestérol régule la fluidité membranaire globale, c'est-à-dire qu'il contrôle la mobilité des différents composants de la membrane cellulaire en fonction de sa concentration. À haute température, le cholestérol inhibe le mouvement des chaînes d'acides gras des phospholipides, ce qui réduit la perméabilité aux petites molécules et la fluidité membranaire. L'inverse est vrai à basse température : la production de cholestérol, et donc sa concentration, est augmentée en réponse au froid. À basse température, le cholestérol interfère avec les interactions des chaînes d'acides gras. Agissant comme un antigel, il maintient la fluidité membranaire. Le cholestérol est plus abondant chez les animaux des régions froides que chez ceux des régions chaudes. Chez les plantes, qui sont dépourvues de cholestérol, d'autres composés stéroliques apparentés remplissent la même fonction.
Phospholipides formant des vésicules lipidiques
Les vésicules lipidiques, ou liposomes, sont des structures approximativement sphériques entourées d'une bicouche lipidique . Elles sont utilisées en laboratoire pour étudier les effets de substances chimiques sur les cellules en les y introduisant directement, et pour mieux comprendre la perméabilité de la membrane cellulaire. Les vésicules lipidiques et les liposomes sont formés en mettant d'abord un lipide en suspension dans une solution aqueuse, puis en agitant le mélange par sonication , ce qui conduit à la formation d'une vésicule. La mesure du flux d' efflux de l'intérieur de la vésicule vers la solution environnante permet aux chercheurs de mieux comprendre la perméabilité membranaire. Les vésicules peuvent être formées avec des molécules et des ions à l'intérieur, en les incorporant à la solution. Des protéines peuvent également être incorporées à la membrane en les solubilisant en présence de détergents et en les fixant aux phospholipides constituant le liposome. Ces structures offrent aux chercheurs un outil pour étudier diverses fonctions des protéines membranaires.des glucides , principalement des glycoprotéines , mais aussi quelques glycolipides ( cérébrosides et gangliosides ). Les glucides jouent un rôle important dans la reconnaissance intercellulaire chez les eucaryotes ; ils sont situés à la surface de la cellule où ils reconnaissent les cellules hôtes et échangent des informations. Les virus qui se lient aux cellules grâce à ces récepteurs provoquent une infection. La glycosylation est généralement absente des membranes intracellulaires ; elle se produit plutôt sur la surface extracellulaire de la membrane plasmique. Le glycocalyx est une structure importante de toutes les cellules, en particulier des épithéliums à microvillosités. Des données récentes suggèrent que le glycocalyx participe à l’adhésion cellulaire, au ciblage des lymphocytes [ à de nombreux autres processus. L’ avant-dernier sucre est le galactose et le sucre terminal est l’acide sialique , car le squelette glucidique est modifié dans l’ appareil de Golgi . L’acide sialique porte une charge négative, formant ainsi une barrière externe aux particules chargées.
Protéines
| Taper | Description | Exemples |
|---|---|---|
| Protéines intégrales ou protéines transmembranaires | Elle traverse la membrane et possède un domaine cytosolique hydrophile interagissant avec les molécules internes, un domaine transmembranaire hydrophobe l'ancrant à la membrane cellulaire, et un domaine extracellulaire hydrophile interagissant avec les molécules externes. Le domaine hydrophobe est constitué d'un ou plusieurs motifs protéiques en hélice α et en feuillet β , ou d'une combinaison de ces motifs . | Canaux ioniques, pompes à protons , récepteurs couplés aux protéines G |
| Protéines ancrées aux lipides | Liées de manière covalente à une ou plusieurs molécules lipidiques, elles s'insèrent par interaction hydrophobe dans la membrane cellulaire et y ancrent la protéine. La protéine elle-même n'est pas en contact avec la membrane. | protéines G |
| Protéines périphériques | Elles sont fixées à des protéines membranaires intégrales ou associées à des régions périphériques de la bicouche lipidique. Ces protéines n'interagissent généralement que temporairement avec les membranes biologiques ; une fois la réaction effectuée, la molécule se dissocie pour poursuivre son action dans le cytoplasme. | Certaines enzymes , certaines hormones |
La membrane cellulaire est riche en protéines, qui représentent généralement environ 50 % de son volume Ces protéines sont essentielles à la cellule car elles sont responsables de diverses activités biologiques. Chez la levure , environ un tiers des gènes codent spécifiquement pour ces protéines, et cette proportion est encore plus élevée chez les organismes multicellulaires . Les protéines membranaires se répartissent en trois grandes catégories : les protéines intégrales, les protéines périphériques et les protéines ancrées aux lipides
Comme indiqué dans le tableau ci-contre, les protéines intégrales sont des protéines transmembranaires amphipathiques. Parmi les exemples de protéines intégrales, on trouve les canaux ioniques, les pompes à protons et les récepteurs couplés aux protéines G. Les canaux ioniques permettent aux ions inorganiques, tels que le sodium, le potassium, le calcium ou le chlore, de diffuser selon leur gradient électrochimique à travers la bicouche lipidique, via des pores hydrophiles perméables à la membrane. L'activité électrique des cellules (par exemple, les cellules nerveuses) est contrôlée par les canaux ioniques. Les pompes à protons sont des pompes protéiques ancrées dans la bicouche lipidique qui permettent le passage des protons à travers la membrane par transfert d'une chaîne latérale d'acide aminé à une autre. Des processus tels que le transport d'électrons et la synthèse d'ATP font appel aux pompes à protons. Un récepteur couplé aux protéines G est une chaîne polypeptidique unique qui traverse la bicouche lipidique sept fois en réponse à des molécules de signalisation (par exemple, des hormones et des neurotransmetteurs). Les récepteurs couplés aux protéines G interviennent dans des processus tels que la communication intercellulaire, la régulation de la production d'AMPc et la régulation des canaux ioniques.
La membrane cellulaire, exposée au milieu extérieur, est un site important de communication intercellulaire. De ce fait, une grande variété de récepteurs protéiques et de protéines d'identification, comme les antigènes , sont présents à sa surface. Les protéines membranaires peuvent également assurer diverses fonctions, telles que le contact intercellulaire, la reconnaissance de surface, l'interaction avec le cytosquelette, la signalisation, l'activité enzymatique ou le transport de substances à travers la membrane.
La plupart des protéines membranaires doivent être insérées dans la membrane. Pour ce faire, une séquence signal N-terminale d'acides aminés dirige les protéines vers le réticulum endoplasmique , qui les insère dans une bicouche lipidique. Une fois insérées, les protéines sont transportées vers leur destination finale dans des vésicules, où celles-ci fusionnent avec la membrane cible.
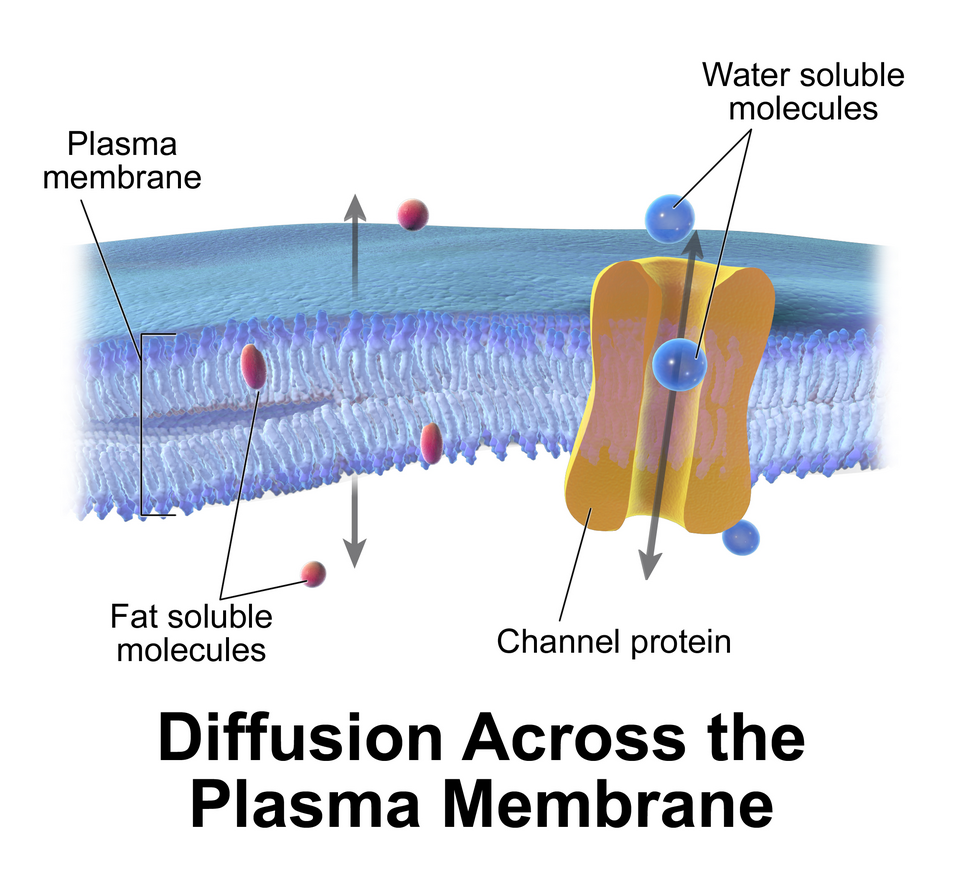
La membrane cellulaire entoure le cytoplasme des cellules vivantes, séparant physiquement les composants intracellulaires du milieu extracellulaire . Elle joue également un rôle dans l'ancrage du cytosquelette , conférant ainsi sa forme à la cellule, et dans l'attachement à la matrice extracellulaire et aux autres cellules, assurant la cohésion des tissus . Les champignons , les bactéries , la plupart des archées et les plantes possèdent également une paroi cellulaire , qui assure un soutien mécanique à la cellule et empêche le passage des grosses molécules .
La membrane cellulaire est sélectivement perméable et capable de réguler les échanges entre l'intérieur et l'extérieur de la cellule, facilitant ainsi le transport des substances nécessaires à sa survie. Le passage des substances à travers la membrane peut s'effectuer soit par transport passif , sans apport d'énergie cellulaire, soit par transport actif , nécessitant un investissement énergétique de la part de la cellule. La membrane maintient également le potentiel membranaire . Elle fonctionne donc comme un filtre sélectif, ne laissant passer que certaines substances. La cellule utilise plusieurs mécanismes de transport impliquant les membranes biologiques.
- Osmose passive et diffusion : Certaines substances (petites molécules, ions), comme le dioxyde de carbone (CO₂ ) et l'oxygène (O₂ ) , peuvent traverser la membrane plasmique par diffusion, un processus de transport passif. La membrane faisant office de barrière pour certaines molécules et certains ions, leurs concentrations peuvent différer de part et d'autre de la membrane. La diffusion se produit lorsque les petites molécules et les ions se déplacent librement d'une zone de forte concentration vers une zone de faible concentration afin d'équilibrer la membrane. Ce processus est considéré comme passif car il ne nécessite pas d'énergie et est propulsé par le gradient de concentration créé de chaque côté de la membrane. Un tel gradient de concentration à travers une membrane semi-perméable induit un flux osmotique pour l'eau. Dans les systèmes biologiques, l'osmose implique le déplacement d'un solvant à travers une membrane semi-perméable, de manière similaire à la diffusion passive, car le solvant se déplace également selon son gradient de concentration et ne requiert aucune énergie. Bien que l'eau soit le solvant le plus courant dans la cellule, il peut également s'agir d'autres liquides, ainsi que de liquides et de gaz supercritiques.
- Canaux et transporteurs protéiques transmembranaires : Les protéines transmembranaires traversent la bicouche lipidique des membranes ; elles fonctionnent de part et d’autre de la membrane pour transporter des molécules. Les nutriments, tels que les sucres ou les acides aminés, doivent pénétrer dans la cellule, et certains produits du métabolisme doivent en sortir. Ces molécules peuvent diffuser passivement à travers des canaux protéiques, comme les aquaporines ( diffusion facilitée) , ou être transportées à travers la membrane par des transporteurs transmembranaires . Les protéines formant des canaux, également appelées perméases , sont généralement très spécifiques et ne reconnaissent et ne transportent qu’une variété limitée de substances chimiques, souvent une seule. Un autre exemple de protéine transmembranaire est un récepteur de surface cellulaire, qui permet la communication entre les cellules par les molécules de signalisation.
- Endocytose : L'endocytose est le processus par lequel les cellules absorbent des molécules en les englobant. La membrane plasmique se déforme légèrement vers l'intérieur, formant une invagination dans laquelle la substance à transporter est capturée. Cette invagination est due à des protéines situées à la surface de la membrane cellulaire, qui agissent comme récepteurs et s'agrègent en dépressions favorisant l'accumulation de protéines et de lipides du côté cytosolique de la membrane. L'invagination se détache ensuite de la membrane à l'intérieur de la cellule, créant une vésicule contenant la substance capturée. L'endocytose est une voie d'internalisation des particules solides (phagocytose ) , des petites molécules et des ions ( pinocytose ) et des macromolécules. L'endocytose nécessite de l'énergie et constitue donc une forme de transport actif.
- Exocytose : De même que des substances peuvent pénétrer dans la cellule par invagination et formation d'une vésicule, la membrane d'une vésicule peut fusionner avec la membrane plasmique, libérant ainsi son contenu dans le milieu environnant. C'est le processus d'exocytose. L'exocytose se produit dans diverses cellules pour éliminer les résidus non digérés de substances internalisées par endocytose, pour sécréter des substances telles que des hormones et des enzymes, et pour transporter une substance à travers une barrière cellulaire. Lors de l'exocytose, la vacuole digestive contenant les déchets non digérés ou la vésicule sécrétoire issue de l'appareil de Golgi est d'abord transportée par le cytosquelette de l'intérieur de la cellule vers sa surface. La membrane de la vésicule entre alors en contact avec la membrane plasmique. Les molécules lipidiques des deux bicouches se réorganisent et les deux membranes fusionnent. Un passage se forme dans la membrane fusionnée et la vésicule libère son contenu hors de la cellule.
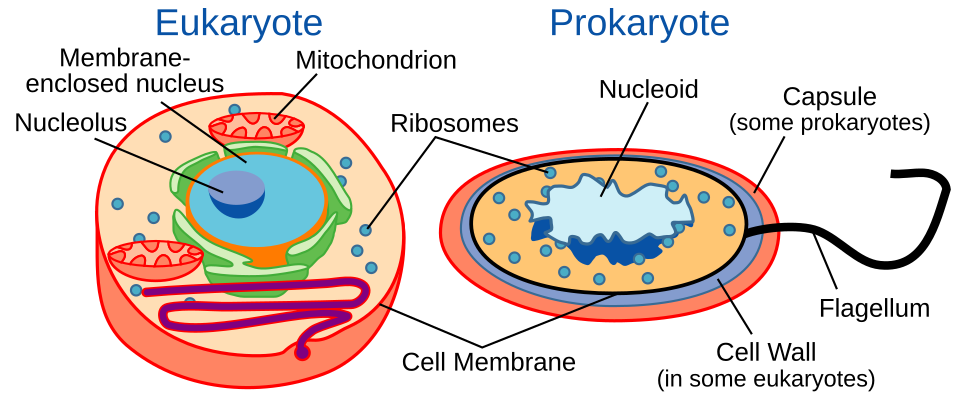
Procaryotes
Les procaryotes sont divisés en deux groupes : les archées et les bactéries. Ces dernières se subdivisent en bactéries Gram-positives et Gram-négatives . Les bactéries Gram-négatives possèdent une membrane plasmique et une membrane externe séparées par le périplasme ; les autres procaryotes , quant à eux, ne possèdent qu’une membrane plasmique. Ces deux membranes diffèrent à plusieurs égards. La membrane externe des bactéries Gram-négatives se distingue de celle des autres procaryotes par la présence de lipopolysaccharides à l’extérieur de la bicouche lipidique, et de lipoprotéines et de phospholipides à l’intérieur . Cette membrane externe est généralement poreuse grâce à des protéines membranaires telles que les porines , qui forment des pores. La membrane plasmique interne est généralement symétrique, tandis que la membrane externe est asymétrique en raison de la présence de protéines comme celles mentionnées précédemment.Les bactéries sont également entourées d'une paroi cellulaire composée de peptidoglycane (acides aminés et sucres). Certaines cellules eucaryotes possèdent aussi une paroi cellulaire, mais aucune n'est constituée de peptidoglycane. La membrane externe des bactéries Gram négatives est riche en lipopolysaccharides, composés de poly- ou oligosaccharides et de saccharolipides , qui stimulent l'immunité naturelle de la cellule. Cette membrane externe peut former des protrusions périplasmiques en conditions de stress ou lors de la rencontre avec une cellule hôte cible, afin de répondre aux besoins de virulence. Ces protrusions peuvent ainsi agir comme des organites de virulence. Les cellules bactériennes offrent de nombreux exemples de la diversité des adaptations des membranes cellulaires procaryotes, avec des structures adaptées à la niche de l'organisme. Par exemple, des protéines présentes à la surface de certaines cellules bactériennes facilitent leur déplacement par glissement . De nombreuses bactéries Gram négatives possèdent des membranes cellulaires contenant des systèmes d'exportation de protéines dépendants de l'ATP .
Structures
Modèle de mosaïque fluide
Selon le modèle de la mosaïque fluide de S.J. Singer et G.L. Nicolson (1972), qui a remplacé le modèle antérieur de Davson et Danielli , les membranes biologiques peuvent être considérées comme un liquide bidimensionnel dans lequel les molécules de lipides et de protéines diffusent plus ou moins facilement. Bien que les bicouches lipidiques qui constituent la base des membranes forment effectivement des liquides bidimensionnels, la membrane plasmique contient également une grande quantité de protéines, qui lui confèrent une structure plus complexe. Parmi ces structures, on peut citer les complexes protéine-protéine, les structures formées par le cytosquelette d'actine et, potentiellement, les radeaux lipidiques .
bicouche lipidique
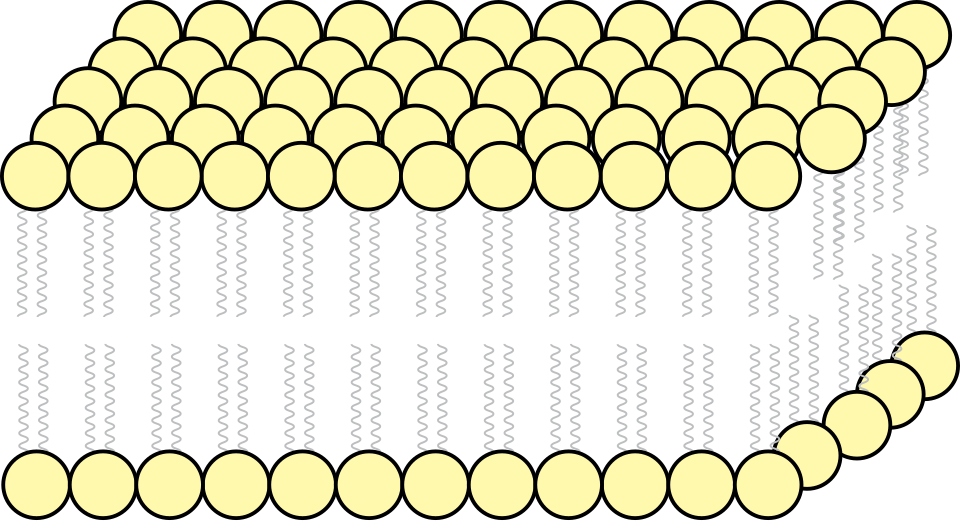
Les bicouches lipidiques se forment par auto-assemblage . La membrane cellulaire est principalement constituée d'une fine couche de phospholipides amphiphiles qui s'organisent spontanément de sorte que les régions hydrophobes, dites « queues », soient isolées de l'eau environnante, tandis que les régions hydrophiles, dites « têtes », interagissent avec les faces intracellulaire (cytosolique) et extracellulaire de la bicouche. Ceci forme une bicouche lipidique sphérique et continue . Les interactions hydrophobes (ou effet hydrophobe ) sont les principales forces motrices de la formation des bicouches lipidiques. L'augmentation des interactions entre les molécules hydrophobes (entraînant le regroupement des régions hydrophobes) permet aux molécules d'eau de se lier plus librement entre elles, augmentant ainsi l'entropie du système. Cette interaction complexe peut inclure des interactions non covalentes telles que les forces de van der Waals , les interactions électrostatiques et les liaisons hydrogène.
Les bicouches lipidiques sont généralement imperméables aux ions et aux molécules polaires. L'agencement des têtes hydrophiles et des queues hydrophobes de la bicouche lipidique empêche la diffusion des solutés polaires (acides aminés, acides nucléiques, glucides, protéines et ions) à travers la membrane, mais permet généralement la diffusion passive des molécules hydrophobes. Ceci confère à la cellule la capacité de contrôler le passage de ces substances via des complexes protéiques transmembranaires tels que les pores, les canaux et les portes. Les flippases et les scramblases concentrent la phosphatidylsérine , porteuse d'une charge négative, sur la face interne de la membrane. Associée à NANA , cette concentration crée une barrière supplémentaire au passage des molécules chargées à travers la membrane.
Les membranes remplissent diverses fonctions dans les cellules eucaryotes et procaryotes . L'un de leurs rôles importants est de réguler les échanges de substances entre l'intérieur et l'extérieur des cellules. La structure en bicouche phospholipidique (modèle de la mosaïque fluide) associée à des protéines membranaires spécifiques explique la perméabilité sélective de la membrane ainsi que les mécanismes de transport passif et actif. De plus, les membranes des procaryotes et celles des mitochondries et des chloroplastes des eucaryotes facilitent la synthèse d'ATP par chimiosmose.
Polarité membranaire
La membrane apicale, ou membrane luminale, d'une cellule polarisée est la surface de la membrane plasmique tournée vers la lumière . Ceci est particulièrement évident dans les cellules épithéliales et endothéliales , mais s'applique également à d'autres cellules polarisées, comme les neurones . La membrane basolatérale, ou membrane basolatérale de la cellule polarisée, est la surface de la membrane plasmique qui forme ses faces basale et latérale . Elle est tournée vers l'extérieur, vers l' interstitium , et opposée à la lumière. Le terme « membrane basolatérale » est un néologisme désignant la membrane basale et la membrane latérale, qui, notamment dans les cellules épithéliales, sont identiques en composition et en activité. Les protéines (comme les canaux ioniques et les pompes ) peuvent librement se déplacer entre la surface basale et la surface latérale de la cellule, et inversement, selon le modèle de la mosaïque fluide . Les jonctions serrées unissent les cellules épithéliales près de leur surface apicale pour empêcher la migration des protéines de la membrane basolatérale vers la membrane apicale. Les surfaces basale et latérale restent ainsi à peu près équivalentes
La membrane cellulaire peut former différents types de structures supramembranaires, telles que les cavéoles , les densités postsynaptiques , les podosomes , les invadopodes , les adhésions focales et différents types de jonctions cellulaires . Ces structures sont généralement responsables de l'adhérence cellulaire , de la communication, de l'endocytose et de l'exocytose . Elles sont composées de protéines spécifiques, comme les intégrines et les cadhérines . Elles peuvent être visualisées par microscopie électronique ou par microscopie à fluorescence .
Cytosquelette
Le cytosquelette, situé sous la membrane cellulaire dans le cytoplasme, sert d'échafaudage aux protéines membranaires pour leur ancrage et permet la formation d'organites qui s'étendent hors de la cellule. En effet, les éléments du cytosquelette interagissent étroitement et de manière extensive avec la membrane cellulaire . Les protéines d'ancrage les confinent à une surface cellulaire particulière – par exemple, la surface apicale des cellules épithéliales qui tapissent l' intestin des vertébrés – et limitent leur diffusion au sein de la bicouche lipidique. Le cytosquelette est capable de former des organites appendiculaires, tels que les cils , extensions microtubulaires recouvertes par la membrane cellulaire, et les filopodes , extensions d' actine . Ces extensions, enveloppées de membrane, font saillie à la surface de la cellule pour percevoir l'environnement extérieur et/ou entrer en contact avec le substrat ou d'autres cellules. Les surfaces apicales des cellules épithéliales sont densément recouvertes de projections digitiformes d'actine appelées microvillosités , qui augmentent la surface cellulaire et, par conséquent, le taux d'absorption des nutriments. Le découplage localisé du cytosquelette et de la membrane cellulaire entraîne la formation d'une vésicule .
Membranes intracellulaires
Le contenu de la cellule, à l'intérieur de sa membrane, est composé de nombreux organites délimités par une membrane , qui contribuent à la fonction globale de la cellule. L'origine, la structure et la fonction de chaque organite engendrent une grande variation dans la composition cellulaire, en raison de la spécificité propre à chaque organite.
- Les mitochondries et les chloroplastes sont considérés comme ayant évolué à partir de bactéries, selon la théorie endosymbiotique . Cette théorie repose sur l'idée que les bactéries Paracoccus et Rhodopseudomonas possèdent des fonctions similaires à celles des mitochondries, et que les cyanobactéries (algues bleues-vertes) possèdent des fonctions similaires à celles des chloroplastes. La théorie endosymbiotique propose qu'au cours de l'évolution, une cellule eucaryote ait phagocyté ces deux types de bactéries, conduisant à la formation de mitochondries et de chloroplastes à l'intérieur de la cellule eucaryote. Cette phagocytose a donné naissance aux systèmes à double membrane de ces organites : la membrane externe provient de la membrane plasmique de l'hôte et la membrane interne de celle de l'endosymbiote. Le fait que les mitochondries et les chloroplastes possèdent leur propre ADN renforce l'hypothèse que ces deux organites ont évolué à partir de bactéries phagocytées qui se sont développées à l'intérieur d'une cellule eucaryote.
- Chez les cellules eucaryotes, la membrane nucléaire sépare le contenu du noyau du cytoplasme. Elle est constituée d'une membrane interne et d' une membrane externe , assurant une régulation stricte des échanges de substances entre le noyau et le cytosol. Ces substances circulent entre le cytosol et le noyau à travers les pores nucléaires . Plus le noyau est actif en transcription , plus sa membrane présente de pores. La composition protéique du noyau peut différer considérablement de celle du cytosol, car de nombreuses protéines ne peuvent pas diffuser à travers ces pores. Au sein de la membrane nucléaire, les membranes interne et externe présentent des compositions protéiques différentes, et seule la membrane externe est en continuité avec la membrane du réticulum endoplasmique (RE). À l'instar du RE, la membrane externe possède également des ribosomes, responsables de la synthèse et du transport des protéines dans l'espace intermembranaire. La membrane nucléaire se désassemble au début de la mitose et se réassemble lors des phases ultérieures.
- Le réticulum endoplasmique (RE), qui fait partie du système endomembranaire, constitue une part importante du contenu membranaire total de la cellule. Le RE est un réseau clos de tubules et de vésicules, dont les principales fonctions sont la synthèse des protéines et le métabolisme des lipides. Il existe deux types de RE : le RE lisse et le RE rugueux. Le RE rugueux possède des ribosomes impliqués dans la synthèse des protéines, tandis que le RE lisse est davantage impliqué dans le traitement des toxines et la régulation du calcium intracellulaire.
- L' appareil de Golgi possède deux citernes golgiennes rondes interconnectées. Ses compartiments forment de multiples réseaux tubulo-réticulaires responsables de l'organisation, de la connexion des cellules et du transport des cargaisons. Ces réseaux présentent des vésicules filiformes continues, semblables à des grappes de raisin, d'une taille comprise entre 50 et 60 nm. L'appareil est constitué de trois compartiments principaux : une citerne plate en forme de disque contenant des réseaux tubulo-réticulaires et des vésicules.
Variations
La membrane cellulaire présente des compositions lipidiques et protéiques différentes selon les types de cellules et peut donc porter des noms spécifiques pour certains types cellulaires.
- Sarcolemme des cellules musculaires : Le sarcolemme est le nom donné à la membrane cellulaire des cellules musculaires. Bien que similaire à d’autres membranes cellulaires, le sarcolemme possède des fonctions qui lui sont propres. Par exemple, il transmet les signaux synaptiques, contribue à la génération des potentiels d’action et joue un rôle essentiel dans la contraction musculaire. Contrairement aux autres membranes cellulaires, le sarcolemme est constitué de petits canaux appelés tubules T qui traversent toute la cellule musculaire. On a également constaté que l’ épaisseur moyenne du sarcolemme est de 10 nm, contre 4 nm pour une membrane cellulaire classique.
- L'oolemme est la membrane cellulaire d'un ovocyte : l'oolemme d'un ovocyte (ovule immature) ne présente pas de bicouche lipidique et n'est pas constitué de lipides . Sa structure comprend une couche interne, l'enveloppe de fécondation, et une couche externe, la zone pellucide (membrane vitelline chez les non-mammifères), composée de glycoprotéines. Des canaux et des protéines sont néanmoins présents pour assurer leurs fonctions membranaires.
- Axolemme : Membrane plasmique spécialisée des axones des cellules nerveuses, responsable de la génération du potentiel d’action. Elle est constituée d’une bicouche lipidique granuleuse et dense, en étroite interaction avec les composants du cytosquelette, la spectrine et l’actine . Ces composants du cytosquelette peuvent se lier aux protéines transmembranaires de l’axolemme et interagir avec elles.
Perméabilité
Plus d articles de Worldlex Wiki
Revenez a l index pour explorer davantage de pages sur l histoire, la science, la culture, la geographie et la societe en francais.
Explorer l index