Les axones sont recouverts d'une membrane appelée axolemme ; le cytoplasme à l'intérieur d'un axone est appelé axoplasme . La plupart des axones se ramifient, parfois abondamment. Les extrémités d'un axone sont appelées télodendrons . L'extrémité renflée d'un télodendron est appelée terminaison axonale , ou pied terminal, et se connecte à la dendrite ou au corps cellulaire d'un autre neurone pour former une synapse . Les axones établissent généralement des contacts avec d'autres neurones au niveau de jonctions appelées synapses , mais ils peuvent également entrer en contact avec des cellules musculaires ou glandulaires. Dans certains cas, l'axone d'un neurone peut former une synapse avec les dendrites de ce même neurone, formant ainsi une autapse . Au niveau d'une synapse, la membrane de l'axone est étroitement accolée à la membrane de la cellule cible, et des structures moléculaires spécifiques permettent la transmission de signaux électriques ou électrochimiques à travers l'espace synaptique. Certaines jonctions synaptiques apparaissent tout au long de l'axone. Ces boutons sont appelés boutons en passant et peuvent se compter par centaines, voire par milliers, le long d'un axone. D'autres synapses apparaissent sous forme de terminaisons aux extrémités des branches axonales.
Un seul axone, avec toutes ses ramifications, peut cibler plusieurs régions du cerveau et générer des milliers de terminaisons synaptiques. Un faisceau d'axones forme un nerveux dans le système nerveux central [ et un fascicule dans le système nerveux périphérique . Chez les mammifères placentaires , le plus grand faisceau de substance blanche du cerveau est le corps calleux , composé d'environ 200 millions d'axones chez l' humain .

Les axones constituent les principales voies de transmission du système nerveux . En faisceaux, ils forment les nerfs du système nerveux périphérique et les voies nerveuses du système nerveux central (SNC). Certains axones peuvent atteindre un mètre, voire plus, tandis que d'autres ne mesurent qu'un millimètre. Les axones les plus longs du corps humain sont ceux du nerf sciatique , qui s'étendent de la base de la moelle épinière jusqu'au gros orteil de chaque pied. Le diamètre des axones est également variable. La plupart des axones individuels ont un diamètre microscopique (généralement d'environ un micromètre ). Les plus grands axones de mammifères peuvent atteindre un diamètre de 20 µm. L' axone géant du calmar , spécialisé dans la conduction très rapide des signaux, mesure près d'un millimètre de diamètre, soit la taille d'une mine de crayon. Le nombre de télodendrons (les ramifications à l'extrémité de l'axone) peut également varier d'une fibre nerveuse à l'autre. Dans le système nerveux central, les axones présentent généralement de multiples télodendrons, avec de nombreuses terminaisons synaptiques. En comparaison, l' axone des cellules granulaires du cervelet est caractérisé par un unique nœud de ramification en forme de T d'où partent deux fibres parallèles . Cette ramification complexe permet la transmission simultanée de messages à un grand nombre de neurones cibles au sein d'une même région cérébrale.
Le système nerveux comprend deux types d'axones : les axones myélinisés et les axones amyéliniques . La myéline est une couche isolante composée d'une substance grasse, formée par deux types de cellules gliales : les cellules de Schwann et les oligodendrocytes . Dans le système nerveux périphérique, les cellules de Schwann forment la gaine de myéline de l'axone myélinisé. Dans le système nerveux central, les oligodendrocytes forment la myéline isolante. Le long des fibres nerveuses myélinisées, des interruptions de la gaine de myéline, appelées nœuds de Ranvier, sont présentes à intervalles réguliers. La myélinisation permet une propagation particulièrement rapide de l'influx nerveux, appelée conduction saltatoire .
Les axones myélinisés des neurones corticaux constituent la majeure partie du tissu nerveux appelé substance blanche dans le cerveau. La myéline confère à ce tissu son aspect blanc , contrairement à la substance grise du cortex cérébral qui contient les corps cellulaires des neurones. Une organisation similaire est observée dans le cervelet . Les faisceaux d'axones myélinisés forment les voies nerveuses du système nerveux central (SNC), et à leurs points de croisement de la ligne médiane du cerveau pour connecter des régions opposées, on les appelle commissures . La plus importante de ces commissures est le corps calleux, qui relie les deux hémisphères cérébraux et contient environ 20 millions d'axones.
La structure d'un neurone se compose de deux régions fonctionnelles distinctes, ou compartiments : le corps cellulaire et les dendrites d'une part, et la région axonale d'autre part.corps de Nissl , qui produisent les protéines neuronales, sont absents de la région axonale. Les protéines nécessaires à la croissance de l'axone et à l'élimination des déchets requièrent un réseau de transport. Ce transport axonal est assuré dans l'axoplasme par des arrangements de microtubules et de filaments intermédiaires de type IV, appelés neurofilaments .
Cône d'émergence de l'axone
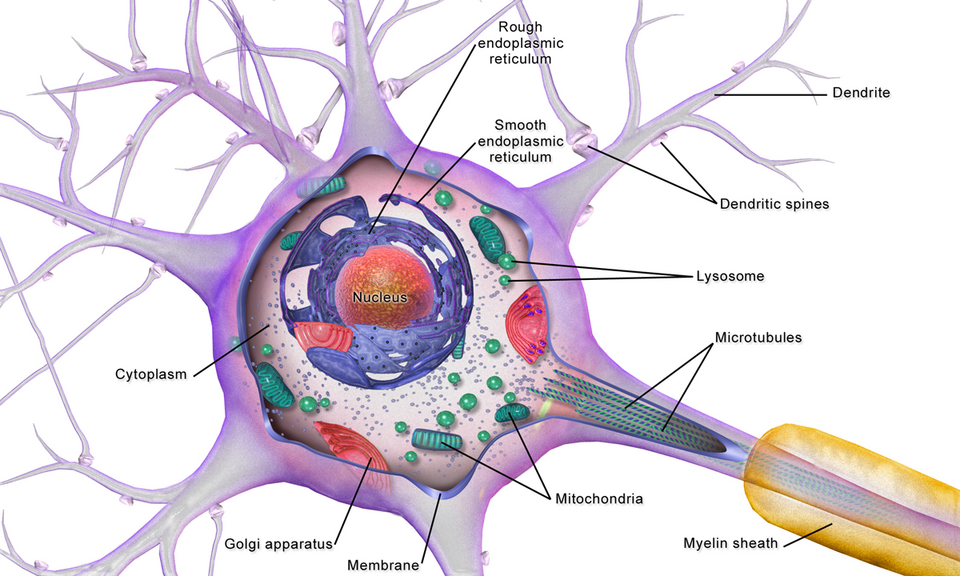
Le cône d'émergence est la zone formée par le corps cellulaire du neurone lors de son prolongement pour former l'axone. Il précède le segment initial. Les potentiels d'action reçus , qui sont sommés dans le neurone, sont transmis au cône d'émergence pour la génération d'un potentiel d'action à partir du segment initial.
Segment initial axonal
Le segment initial de l'axone (SIA) est un microdomaine de l'axone structurellement et fonctionnellement distinct. L'une de ses fonctions est de séparer la partie principale de l'axone du reste du neurone ; une autre est de contribuer à l'initiation des potentiels d'action. Ces deux fonctions soutiennent la polarité cellulaire neuronale : les dendrites (et, dans certains cas, le soma ) reçoivent les signaux d'entrée au niveau de la région basale, tandis que l'axone du neurone émet les signaux de sortie au niveau de la région apicale.
Le segment initial de l'axone est amyélinisé et contient un complexe protéique spécialisé. Sa longueur est comprise entre 20 et 60 μm environ et il constitue le site d'initiation du potentiel d'action. La position et la longueur du segment initial de l'axone peuvent varier, témoignant d'une plasticité permettant d'ajuster finement l'activité neuronale. Un segment initial de l'axone plus long est associé à une excitabilité accrue. Cette plasticité se manifeste également par la capacité du segment initial de l'axone à modifier sa distribution et à maintenir l'activité des circuits neuronaux à un niveau constant.
Le segment initial de l'axone (SIA) est hautement spécialisé pour la conduction rapide de l'influx nerveux . Ceci est rendu possible par une forte concentration de canaux sodiques voltage-dépendants dans le segment initial où le potentiel d'action est initié. Ces canaux ioniques sont associés à un grand nombre de molécules d'adhérence cellulaire et de protéines d'échafaudage qui les ancrent au cytosquelette. Les interactions avec l'ankyrine-G sont importantes car cette protéine est le principal organisateur du SIA.
Dans d'autres cas, comme observé chez le rat, un axone prend naissance à partir d'une dendrite ; on dit alors que ces axones sont d'origine dendritique. Certains axones d'origine dendritique possèdent un segment initial proximal, qui débute directement à l'origine de l'axone, tandis que d'autres possèdent un segment initial distal, nettement séparé de l'origine de l'axone. Chez de nombreuses espèces, certains neurones possèdent des axones qui émanent de la dendrite et non du corps cellulaire ; on les appelle des dendrites porteuses d'axones. Dans de nombreux cas, un axone prend naissance au niveau d'un cône d'émergence sur le soma ; on dit alors que ces axones sont d'origine somatique. Certains axones d'origine somatique possèdent un segment initial proximal, adjacent au cône d'émergence, tandis que d'autres possèdent un segment initial distal, séparé du soma par un cône d'émergence prolongé.
Transport axonal
Le transport antérograde sortant du corps cellulaire, le long de l'axone, achemine les mitochondries et les protéines membranaires nécessaires à la croissance jusqu'à la terminaison axonale. Le transport rétrograde entrant transporte les déchets cellulaires de la terminaison axonale vers le corps cellulaire. Les voies de transport sortant et entrant utilisent différents ensembles de protéines motrices . Le transport sortant est assuré par la kinésine , et le transport entrant par la dynéine . La dynéine se déplace vers l'extrémité moins de l'axone. Il existe de nombreuses isoformes de kinésine et de dynéine, et chacune transporterait une cargaison différente. Les études sur le transport dans l'axone ont conduit à la dénomination de la kinésine.
Myélinisation
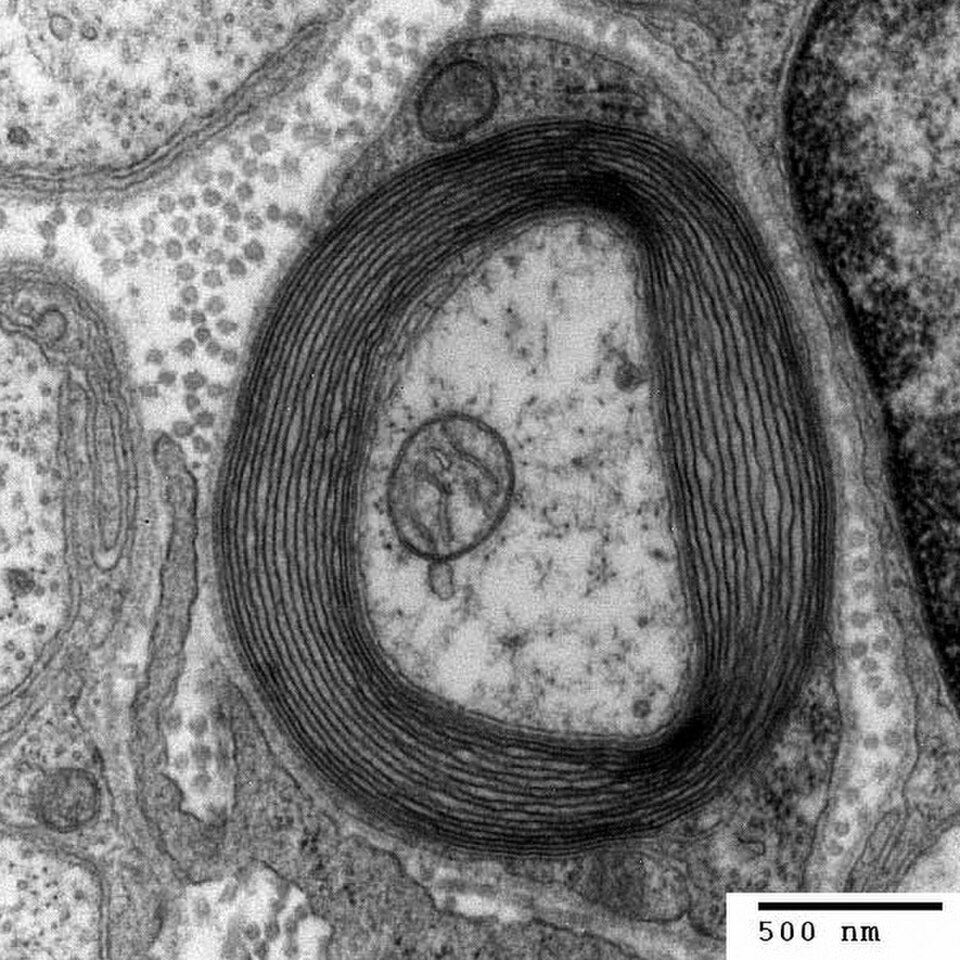
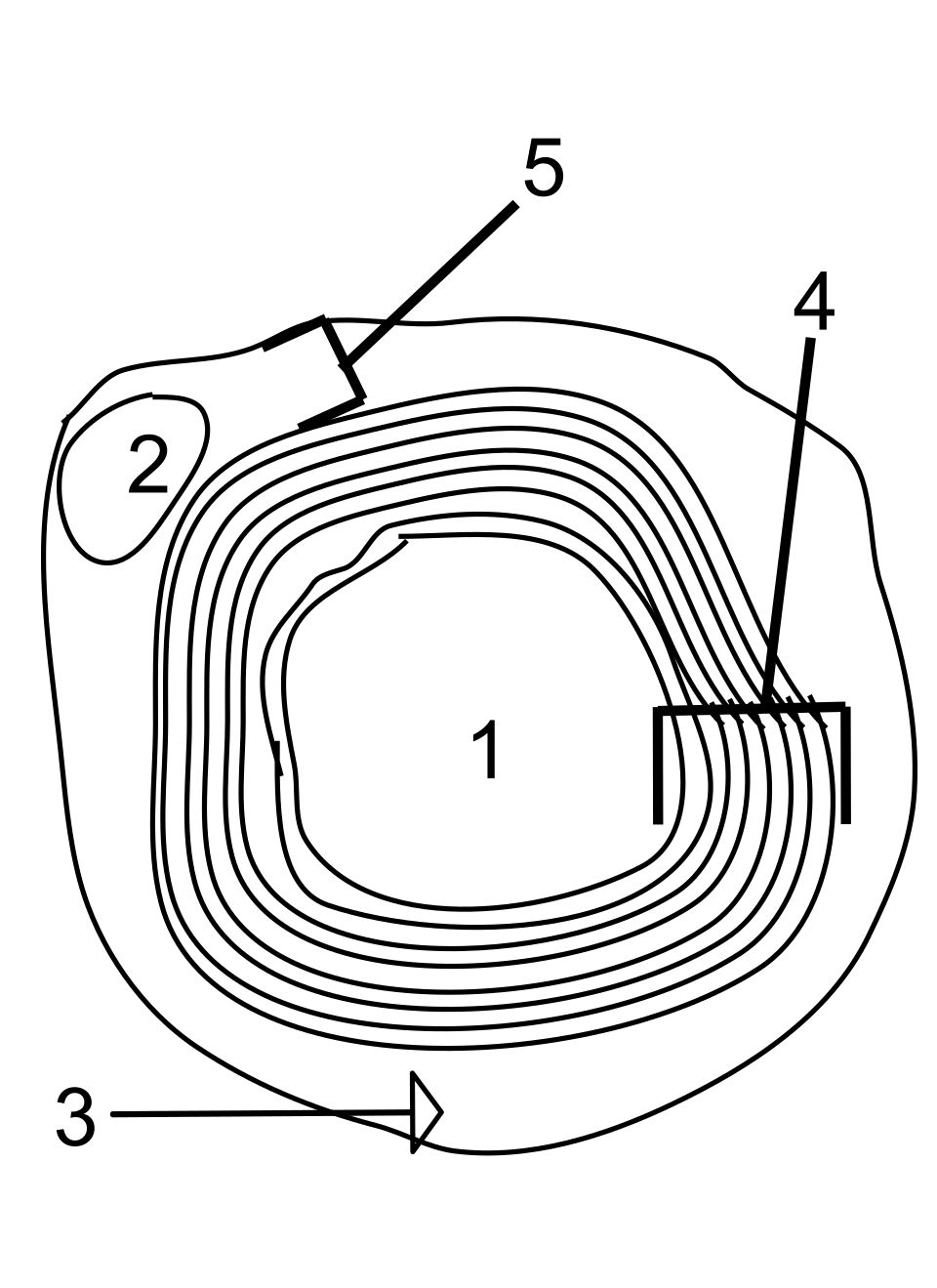
Dans le système nerveux, les axones peuvent être myélinisés ou non myélinisés. La myélinisation consiste en la présence d'une couche isolante appelée gaine de myéline. La membrane de myéline est unique par son rapport lipides/protéines relativement élevé.
Dans le système nerveux périphérique, les axones sont myélinisés par des cellules gliales appelées cellules de Schwann . Dans le système nerveux central, la gaine de myéline est fournie par un autre type de cellule gliale, l' oligodendrocyte . Les cellules de Schwann myélinisent un seul axone. Un oligodendrocyte peut myéliniser jusqu'à 50 axones.
La composition de la myéline diffère entre les deux types de systèmes. Dans le SNC, la protéine myélinique majoritaire est la protéine protéolipidique , et dans le SNP, il s'agit de la protéine basique de la myéline .
Nœuds de Ranvier
Terminaisons axonales
Varices axonales
Dans un cerveau au développement normal, des boutons présynaptiques, également appelés varicosités axonales , sont présents le long de certains axones. On les retrouve notamment dans les régions de l' hippocampe impliquées dans la libération des neurotransmetteurs . Cependant, les varicosités axonales sont également présentes dans les maladies neurodégénératives, où elles perturbent la conduction du potentiel d'action. Elles constituent aussi une caractéristique des traumatismes crâniens . Les lésions axonales affectent généralement le cytosquelette de l'axone, perturbant ainsi le transport des neurotransmetteurs. Il peut en résulter une accumulation de protéines, comme le précurseur de la protéine bêta-amyloïde, formant un gonflement et de nombreuses varicosités le long de l'axone
Potentiels d'action
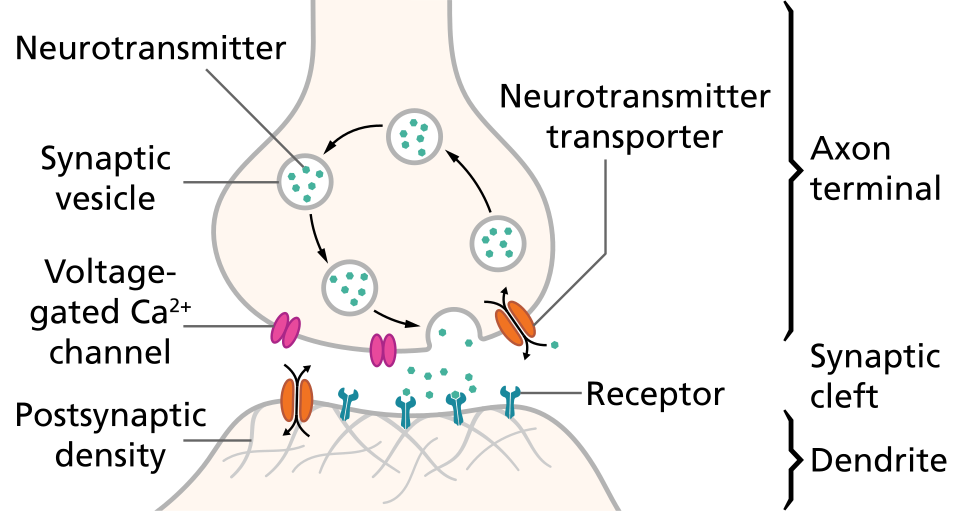
La plupart des axones transmettent des signaux sous forme de potentiels d'action, qui sont des impulsions électrochimiques discrètes se propageant rapidement le long de l'axone, depuis le corps cellulaire jusqu'aux points de contact synaptique avec les cellules cibles. La caractéristique fondamentale d'un potentiel d'action est son caractère « tout ou rien » : chaque potentiel d'action généré par un axone possède sensiblement la même amplitude et la même forme. Cette caractéristique permet la transmission des potentiels d'action d'une extrémité à l'autre d'un long axone sans aucune atténuation. Il existe cependant certains types de neurones à axones courts qui transmettent des signaux électrochimiques gradués, d'amplitude variable.
Lorsqu'un potentiel d'action atteint une terminaison présynaptique, il active la transmission synaptique. La première étape est l'ouverture rapide des canaux calciques de la membrane de l'axone, permettant aux ions calcium de pénétrer dans la cellule. L'augmentation de la concentration intracellulaire de calcium qui en résulte provoque la fusion des vésicules synaptiques (minuscules structures entourées d'une membrane lipidique) remplies de neurotransmetteur avec la membrane de l'axone et la libération de leur contenu dans l'espace extracellulaire. Le neurotransmetteur est libéré du neurone présynaptique par exocytose . Il diffuse ensuite jusqu'aux récepteurs situés sur la membrane de la cellule cible. Le neurotransmetteur se lie à ces récepteurs et les active. Selon le type de récepteurs activés, l'effet sur la cellule cible peut être l'excitation, l'inhibition ou une modification de son métabolisme. Cette séquence d'événements se déroule généralement en moins d'un millième de seconde. Ensuite, à l'intérieur de la terminaison présynaptique, un nouvel ensemble de vésicules se positionne contre la membrane, prêt à être libéré lors du prochain potentiel d'action. Le potentiel d'action constitue l'étape électrique finale de l'intégration des messages synaptiques à l'échelle du neurone.
L'enregistrement extracellulaire de la propagation des potentiels d'action dans les axones a été démontré chez des animaux en liberté de mouvement. Si les potentiels d'action somatiques extracellulaires ont été utilisés pour étudier l'activité cellulaire chez ces animaux, notamment celle des cellules de lieu , l'activité axonale peut également être enregistrée dans la substance blanche et la substance grise . L'enregistrement extracellulaire de la propagation des potentiels d'action axonaux se distingue des potentiels d'action somatiques par trois caractéristiques : 1. Le signal présente une durée crête-creux plus courte (environ 150 µs) que celle des cellules pyramidales (environ 500 µs) ou des interneurones (environ 250 µs). 2. La variation de potentiel est triphasique. 3. L'activité enregistrée sur une tétrode n'est visible que sur l'un des quatre fils d'enregistrement. Chez des rats en liberté de mouvement, des signaux axonaux ont été isolés dans les faisceaux de substance blanche, notamment l'alvéus et le corps calleux, ainsi que dans la substance grise de l'hippocampe.
En réalité, la génération des potentiels d'action in vivo est de nature séquentielle, et ces pics successifs constituent le code numérique des neurones. Bien que des études antérieures indiquent une origine axonale d'un pic unique évoqué par des impulsions de courte durée, les signaux physiologiques in vivo déclenchent l'initiation de pics séquentiels au niveau des corps cellulaires des neurones.
Outre la propagation des potentiels d'action jusqu'aux terminaisons axonales, l'axone est capable de les amplifier, assurant ainsi une propagation fiable des potentiels d'action successifs vers la terminaison axonale. Au niveau moléculaire, les canaux sodiques voltage-dépendants des axones présentent un seuil d'activation plus bas et une période réfractaire plus courte en réponse à des impulsions de courte durée.
Développement et croissance
Développement
Le développement de l'axone jusqu'à sa cible est l'une des six étapes majeures du développement global du système nerveux . On ignore si la spécification élongation axonale ou inversement , bien que des données récentes penchent pour la seconde hypothèse. Si un axone incomplet est sectionné, sa polarité peut s'inverser et d'autres neurites peuvent potentiellement devenir l'axone. Cette inversion de polarité n'a lieu que lorsque l'axone est sectionné à une longueur d'au moins 10 μm inférieure à celle des autres neurites. Après la section, le neurite le plus long deviendra le futur axone et tous les autres neurites, y compris l'axone initial, se différencieront en dendrites. L'application d'une force externe sur un neurite, provoquant son allongement, le transformera en axone. Néanmoins, le développement axonal est réalisé grâce à une interaction complexe entre la signalisation extracellulaire, la signalisation intracellulaire et la dynamique du cytosquelette .
Signalisation extracellulaire
Les signaux extracellulaires qui se propagent à travers la matrice extracellulaire entourant les neurones jouent un rôle prépondérant dans le développement axonal. Ces molécules de signalisation comprennent des protéines, des facteurs neurotrophiques , ainsi que des molécules de la matrice extracellulaire et d'adhérence. La nétrine (également connue sous le nom d'UNC-6), une protéine sécrétée, intervient dans la formation des axones. Lorsqu'un récepteur de la nétrine, UNC-5, est muté, plusieurs neurites se projettent de manière irrégulière hors des neurones et, finalement, un seul axone se prolonge antérieurement. Les facteurs neurotrophiques – le facteur de croissance nerveuse (NGF), le facteur neurotrophique dérivé du cerveau (BDNF) et la neurotrophine-3 (NTF3) – sont également impliqués dans le développement axonal et se lient aux récepteurs Trk .
L’ enzyme de conversion des gangliosides , la sialidase des gangliosides de la membrane plasmique (PMGS), impliquée dans l’activation de TrkA à l’extrémité des neutrites, est nécessaire à l’élongation des axones. La PMGS se distribue de manière asymétrique à l’extrémité du neurite destiné à devenir le futur axone.
Signalisation intracellulaire
Au cours du développement axonal, l'activité de la PI3K augmente à l'extrémité de l'axone en formation. L'inhibition de l'activité de la PI3K bloque le développement axonal. L'activation de la PI3K entraîne la production de phosphatidylinositol (3,4,5)-trisphosphate (PtdIns), ce qui peut provoquer un allongement significatif du neurite et sa conversion en axone. Ainsi, la surexpression des phosphatases déphosphorylant le PtdIns conduit à un défaut de polarisation.
Dynamique du cytosquelette
Le neurite présentant la plus faible teneur en filaments d'actine deviendra l'axone. La concentration de PGMS et la teneur en actine F sont inversement corrélées ; lorsque le PGMS s'accumule à l'extrémité d'un neurite, sa teneur en actine F diminue considérablement . De plus, l'exposition à des agents dépolymérisants de l'actine et à la toxine B (qui inactive la voie de signalisation Rho ) induit la formation de plusieurs axones. Par conséquent, l'interruption du réseau d'actine dans un cône de croissance favorise la différenciation de son neurite en axone
Croissance
Les axones en croissance se déplacent dans leur environnement grâce au cône de croissance , situé à leur extrémité. Ce cône possède une large extension lamellaire appelée lamellipode , qui contient des protubérances appelées filopodes . Les filopodes permettent à l'axone d'adhérer aux surfaces et d'explorer son environnement. L'actine joue un rôle majeur dans la mobilité de ce système. Les environnements riches en molécules d'adhérence cellulaire (CAM) créent un milieu idéal pour la croissance axonale. Ces molécules semblent fournir une surface « collante » sur laquelle les axones peuvent se développer. Parmi les CAM spécifiques au système nerveux, on trouve N-CAM , TAG-1 ( une glycoprotéine axonale ) et MAG , toutes appartenant à la superfamille des immunoglobulines . Un autre ensemble de molécules, appelées molécules d'adhérence de la matrice extracellulaire , fournit également un substrat collant pour la croissance des axones. La laminine , la fibronectine , la ténascine et le perlécan en sont des exemples . Certains de ces composés se lient à la surface des cellules et agissent donc comme attractifs ou répulsifs à courte portée. D'autres sont des ligands diffusibles et peuvent ainsi avoir des effets à longue portée.
Des cellules appelées cellules guides participent à la croissance des axones neuronaux. Ces cellules, qui contribuent au guidage axonal , sont généralement d'autres neurones, parfois immatures. Une fois la croissance de l'axone achevée au niveau de sa connexion à la cible, son diamètre peut être multiplié par cinq, selon la vitesse de conduction requise.
Des recherches ont également montré que si les axones d'un neurone étaient endommagés, tant que le soma (le corps cellulaire du neurone) ne l'était pas, les axones se régénéraient et recréaient des connexions synaptiques avec les neurones grâce aux cellules guides . Ce phénomène est également appelé neurogénération .
Nogo-A est un composant inhibiteur de la croissance des neurites présent dans les membranes de myéline du système nerveux central (au niveau des axones). Il joue un rôle crucial dans la limitation de la régénération axonale chez les mammifères adultes. Des études récentes ont montré que le blocage et la neutralisation de Nogo-A permettent d'induire une régénération axonale à longue distance, améliorant ainsi la récupération fonctionnelle de la moelle épinière chez le rat et la souris. Ces résultats restent à confirmer chez l'humain. Une étude récente a également mis en évidence que les macrophages activés par une voie inflammatoire spécifique, elle-même activée par le récepteur Dectin-1 , sont capables de favoriser la récupération axonale, tout en induisant une neurotoxicité neuronale.
Réglementation de la longueur
La longueur des axones varie considérablement, de quelques micromètres à plusieurs mètres chez certains animaux. Ceci souligne l'existence d'un mécanisme de régulation de la longueur cellulaire permettant aux neurones de percevoir la longueur de leurs axones et d'en contrôler la croissance. Il a été découvert que les protéines motrices jouent un rôle important dans la régulation de la longueur des axones . À partir de cette observation, des chercheurs ont élaboré un modèle explicite de croissance axonale décrivant comment les protéines motrices peuvent influencer la longueur de l'axone au niveau moléculaire Ces études suggèrent que les protéines motrices transportent des molécules de signalisation du soma au cône de croissance et inversement, dont la concentration oscille au cours du temps à une fréquence dépendant de la longueur de l'axone.
Classification
Les axones sont classés selon deux systèmes. Le premier, introduit par Erlanger et Gasser, regroupe les fibres en trois groupes principaux, désignés par les lettres A, B et C. Ces groupes comprennent à la fois les fibres sensitives ( afférentes ) et les fibres motrices ( efférentes ). Le groupe A est subdivisé en fibres alpha (Aα), bêta (Aβ), gamma (Aγ) et delta ( Aδ). Les motoneurones associés à ces différentes fibres motrices sont les motoneurones inférieurs : les motoneurones alpha , bêta et gamma correspondent respectivement aux fibres nerveuses Aα, Aβ et Aγ.
Des travaux ultérieurs menés par d'autres chercheurs ont identifié deux groupes de fibres Aa sensorielles. Celles-ci ont ensuite été intégrées à une classification (classification de Lloyd) ne regroupant que les fibres sensorielles (bien que certaines d'entre elles soient mixtes et possèdent également des fonctions motrices). Cette classification désigne les groupes de fibres sensorielles par des types et utilise des chiffres romains : type Ia, type Ib, type II, type III et type IV.
Moteur
Les motoneurones inférieurs possèdent deux types de fibres :
| Taper | Classification d' Erlanger-Gasser | Diamètre (μm) | Myéline | Vitesse de conduction (mètres/seconde) | Fibres musculaires associées |
|---|---|---|---|---|---|
| motoneurone alpha (α) | Aα | 13–20 | Oui | 80–120 | Fibres musculaires extrafusales |
| motoneurone bêta (β) | Aβ | ||||
| motoneurone gamma (γ) | Aγ | 5-8 | Oui | 4–24 | Fibres musculaires intrafusales |
Sensoriel
Différents types de fibres nerveuses innervent différents récepteurs sensoriels . Les propriocepteurs sont innervés par les fibres sensorielles de type Ia, Ib et II, les mécanorécepteurs par les fibres sensorielles de type II et III, et les nocicepteurs et thermorécepteurs par les fibres sensorielles de type III et IV.
| Taper | Classification d' Erlanger-Gasser | Diamètre (μm) | Myéline | Vitesse de conduction (m/s) | Récepteurs sensoriels associés | propriocepteurs | mécanorécepteurs | Nocicepteurs et thermorécepteurs |
|---|---|---|---|---|---|---|---|---|
| Je | Aα | 13–20 | Oui | 80–120 | Récepteurs primaires du fuseau neuromusculaire (extrémité annulaire spiralée) | ✔ | ||
| Ib | Aα | 13–20 | Oui | 80–120 | Organe tendineux de Golgi | |||
| II | Aβ | 6–12 | Oui | 33–75 | Récepteurs secondaires du fuseau neuromusculaire (extrémité en forme de fleur). Tous les mécanorécepteurs cutanés. | ✔ | ||
| III | Aδ | 1–5 | Mince | 3–30 | Terminaisons nerveuses libres du toucher et de la pression Nocicepteurs du faisceau spinothalamique latéral Thermorécepteurs du froid | ✔ | ||
| IV | C | 0,2–1,5 | Non | 0,5–2,0 | Nocicepteurs du faisceau spinothalamique antérieur Récepteurs de chaleur |
Autonome
Le système nerveux autonome possède deux types de fibres périphériques :
| Taper | Classification d' Erlanger-Gasser | Diamètre (μm) | Myéline | Vitesse de conduction (m/s) |
|---|---|---|---|---|
| fibres préganglionnaires | B | 0,5–3 | Oui | 3–15 |
| fibres postganglionnaires | C | 0,2–1,4 | Non | 0,5–2,0 |
Signification clinique
Lorsqu'un axone est écrasé, un processus actif de dégénérescence axonale se produit dans la partie de l'axone la plus éloignée du corps cellulaire. Cette dégénérescence survient rapidement après la lésion : la partie de l'axone concernée est obstruée au niveau des membranes et dégradée par les macrophages. Ce phénomène est connu sous le nom de dégénérescence wallérienne . La mort axonale peut également se produire dans de nombreuses maladies neurodégénératives , notamment lorsque le transport axonal est altéré ; on parle alors de dégénérescence de type wallérien . Des études suggèrent que cette dégénérescence résulte de l' incapacité de la protéine axonale NMNAT2 à atteindre l'intégralité de l'axone
La démyélinisation des axones provoque la multitude de symptômes neurologiques observés dans la sclérose en plaques .
La dysmyélinisation est une formation anormale de la gaine de myéline. Elle est impliquée dans plusieurs leucodystrophies , ainsi que dans la schizophrénie .
Un traumatisme crânien grave peut entraîner des lésions étendues des voies nerveuses, endommageant les axones ; on parle alors de lésion axonale diffuse . Ceci peut conduire à un état végétatif persistant . Des études menées chez le rat ont montré que les lésions axonales consécutives à un seul traumatisme crânien léger peuvent engendrer une susceptibilité à de nouvelles lésions après des traumatismes crâniens légers répétés.
Un conduit de guidage nerveux est un moyen artificiel de guider la croissance axonale pour permettre la neurogénération , et constitue l'un des nombreux traitements utilisés pour différents types de lésions nerveuses .
Terminologie
Certains dictionnaires généralistes définissent la « fibre nerveuse » comme tout prolongement neuronal , incluant à la fois les axones et les dendrites . Cependant, les sources médicales utilisent généralement le terme « fibre nerveuse » pour désigner uniquement l’axone.
Histoire
L'anatomiste allemand Otto Friedrich Karl Deiters est généralement reconnu pour avoir découvert l'axone en le distinguant des dendrites. Le Suisse Rüdolf Albert von Kölliker et l'Allemand Robert Remak furent les premiers à identifier et à caractériser le segment initial de l'axone. Kölliker nomma l'axone en 1896. Louis-Antoine Ranvier fut le premier à décrire les brèches ou nœuds présents sur les axones ; ces structures axonales sont aujourd'hui communément appelées nœuds de Ranvier . Santiago Ramón y Cajal , anatomiste espagnol, proposa que les axones constituent les structures de sortie des neurones, et décrivit leur fonction. Joseph Erlanger et Herbert Gasser ont précédemment développé le système de classification des fibres nerveuses périphériques , basé sur la vitesse de conduction axonale, la myélinisation , la taille des fibres, etc. Alan Hodgkin et Andrew Huxley ont également utilisé l'axone géant du calmar (1939) et, dès 1952, ils avaient obtenu une description quantitative complète de la base ionique du potentiel d'action, ce qui a conduit à la formulation du modèle de Hodgkin-Huxley . Hodgkin et Huxley ont reçu conjointement le prix Nobel pour ces travaux en 1963. Les formules détaillant la conductance axonale ont été étendues aux vertébrés dans les équations de Frankenhaeuser-Huxley. La compréhension de la base biochimique de la propagation du potentiel d'action a encore progressé et inclut de nombreux détails sur les canaux ioniques individuels .
Autres animaux
Les axones des invertébrés ont fait l'objet de nombreuses études. Le calmar côtier à longues nageoires , souvent utilisé comme organisme modèle , possède l'axone le plus long connu . Le calmar géant possède l'axone le plus grand connu. Son diamètre varie généralement de 0,5 à 1 mm et il intervient dans le contrôle de son système de propulsion par jet . La vitesse de conduction la plus rapide enregistrée, de 210 m/s, a été observée dans les axones myélinisés de certaines crevettes pélagiques pénéides , et la vitesse habituelle se situe entre 90 et 200 m/s ( contre 100 à 120 m/s pour l'axone myélinisé le plus rapide chez les vertébrés).
Images supplémentaires
